|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: Great Spotted Cuckoo, activity, home range, behaviour.
Actividad No hay datos.
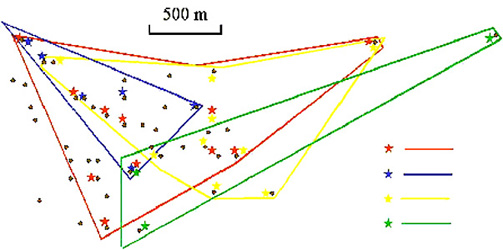
Dominio vital y territorio Debido a que la mayoría de los nidos en los que se ha detectado la muerte de pollos de Críalo eran nidos multiparasitados por más de una hembra (Soler y Soler, 1991; Arias de Reyna, 1998), el comportamiento territorial en esta especie sería adaptativo. Es decir, cada pareja (en el caso de que fueran monógamos) o grupo reproductor, debería de defender un territorio que incluyera un número de nidos de hospedador suficiente para su reproducción (Arias de Reyna, 1998). De acuerdo con esta premisa se han detectado comportamientos territoriales en el Críalo (Mundy y Cook, 1977; Arias de Reyna et al., 1982; Arias de Reyna et al., 1987), estableciéndose el territorio defendido entre 1 km2 (Valverde, 1971), 2,5 (Mundy y Cook, 1977), 2,9 (Arias de Reyna, 1998) y 8-9 km2 (Mountfort y Ferguson-Lees, 1961). Parece que defienden los territorios hasta que no quedan nidos disponibles de hospedador para parasitar y, a partir de ese momento, los críalos dejan de ser territoriales. Esto explicaría el hecho de que frecuentemente distintas hembras parasiten el mismo nido de urraca (Arias de Reyna, 1998). Sin embargo, en otro estudio (Gaston, 1976), no encontraron evidencias de territorialidad en esta especie. Todos estos estudios fueron realizados con individuos sin marcar y, por tanto, los resultados deben de ser analizados con cautela. Por otro lado, un estudio de la distribución espacial y temporal de la puesta de huevos de distintas hembras de Críalo, utilizando marcadores genéticos (Martínez et al.,1998a), reveló un gran solapamiento entre los territorios de las distintas hembras (ver Figura 1), por lo que la territorialidad de esta especie no parece ser importante. De diez nidos multiparasitados (58,3 % de los nidos) en los que se pudieron identificar los padres, cuatro de ellos tenían huevos de más de una hembra y, en los seis nidos multiparasitados restantes, los huevos eran de la misma hembra (Martínez et al., 1998a). El resultado más importante es que la distancia media entre nidos parasitados por la misma hembra era igual o mayor que la distancia media entre nidos parasitados por esa hembra y nidos parasitados por otras. Estos resultados, por tanto, no están de acuerdo con la hipótesis de una territorialidad en hembras.
Figura 1. Localización de los nidos de urraca (puntos negros) y parasitación de esos nidos por 4 hembras (estrellas de distintos colores). También se muestran los territorios establecidos para cada una de esas hembras. Figura modificada de Martínez et al. (1998).
Además, debido a que el sistema de emparejamiento no era poligínico (ver más adelante), tampoco puede haber territorialidad en los machos. Una posibilidad que explicaría estos resultados bajo el prisma de la territorialidad es que ésta fuera temporal y dinámica defendiendo pequeños territorios con unos pocos nidos de hospedador activos que fueran cambiando a lo largo de la estación reproductora. Sin embargo, la distancia espacial entre huevos puestos consecutivamente por la misma hembra resultó ser similar a la distancia media entre huevos puestos por distintas hembras el mismo día, incluso después de tener en cuenta el efecto de la variación en la disponibilidad de nidos. Por tanto, se puede concluir que en el Críalo no existen territorios exclusivos de parasitación (Martínez et al., 1998a). Con respecto al sistema de emparejamiento, utilizando marcadores genéticos se identificaron 4 hembras distintas parasitando nidos de Urraca en el área de estudio (Guadix, Granada). Los huevos puestos por tres de estas hembras habían sido fecundados por dos machos distintos cada una, pero no se detectó que un mismo macho fecundara huevos de más de una hembra. Por tanto, el sistema de emparejamiento puede ser monógamo con algunas cópulas extra-pareja, poliándrico o promiscuo, pero no poligínico (Martínez et al., 1998b). Sin embargo, se identificaron algunos machos poligínicos (Martínez et al., 1998b). Nueve hembras fueron fertilizadas por un macho y dos fueron fertilizados por dos machos. Con respecto a machos, 10 fertilizaron a sólo una hembra y tres fertilizaron a dos. Además, 34 de los 57 pollos (59,6%) para los que se estableció el padre y la madre procedían de parejas genéticamente monógamas, mientras que el resto de pollos procedían de fertilizaciones en las que el padre y/o la madre era polígamo. Aunque estos datos indican que la mayoría de las parejas son monógamas, y que algunos individuos son polígamos, la existencia de mecanismos de competencia espermática no nos permiten concluir que la estrategia de emparejamiento social más común en el Críalo sea la monogamia. Además pudo ocurrir que algunos de los individuos clasificados como monógamos en los análisis de paternidad podrían haber estado involucrados en cópulas "extra-pareja" y ninguna de estas cópulas fuera efectiva. Por tanto, el porcentaje de parejas monógamos establecida por medio de análisis de paternidad debería de ser interpretada como el límite superior de monogamia (Martínez et al., 1998b). Es decir, se necesitan datos de comportamiento social para establecer más precisamente las relaciones sociales existentes entre machos y hembras de Críalo.
Impronta, patrones sociales y territorialidad ¿Cómo reconocen los críalos jóvenes la especie a la que pertenecen? La mayoría de las especies de aves, durante los primeros días de vida, aprenden la apariencia de sus padres, lo que posteriormente les permite reconocer a los individuos de su propia especie. Cuando las aves son criadas por otras especies, esta impronta tiene consecuencias inmediatas en el comportamiento filial y social (impronta filial) y, cuando llegan a adultos, intentan emparejarse con individuos de la especie que los ha criado y no con individuos de su propia especie (impronta sexual; Lorenz, 1935). Un aspecto relevante es que el contacto con hermanos o jóvenes de la misma especie es muy importante para establecer las preferencias sociales por individuos de su propia especie (Lorenz, 1935; Kruijt et al., 1983). De hecho, se ha sugerido la posibilidad de la existencia de un retraso en el periodo dedicado al aprendizaje de la apariencia de los individuos de su propia especie hasta una edad en la que la apariencia de los jóvenes sea muy parecida a la de los adultos (Bateson, 1979). Las especies parásitas de cría obligadas no tienen contactos con individuos adultos o, en el caso de que existan, son muy esporádicos, por lo que el reconocimiento y emparejamiento con individuos de la misma especie podría ser una característica innata (Tinbergen, 1951). Otra posibilidad sería que los contactos existentes entre jóvenes de la misma especie parásita fueran suficientes para conseguir un aprendizaje de las características propias de la especie que, a su vez, explicaran el posterior emparejamiento (Freeberg et al., 1995; Soler y Soler, 1999). En este sentido, la mayoría (63%) de los nidos de Urraca parasitados por el Críalo en la Hoya de Guadix reciben más de un huevo parásito, por lo que en la mayoría de los nidos conviven más de un pollo parásito. Además, los pollos de críalo al abandonar el nido permanecen en la zona de parasitación durante varios meses, por lo que la probabilidad de que individuos jóvenes procedentes de distintos nidos de hospedador se encuentren es elevada. Estas dos características hacen que la probabilidad de contacto entre jóvenes críalos sea prácticamente del 100%. De hecho ya se ha mencionado que jóvenes críalos procedentes de distintos nidos se agregan formando grupos de jóvenes parásitos que son alimentados por urracas adultas (Soler et al., 1995; sólo dos de 62 jóvenes seguidos no llegaron a formar parte de ningún grupo de críalos). Por otro lado, los pollos y jóvenes de Críalo reciben visitas esporádicas de críalos adultos y, estas visitas, podrían servir para que los jóvenes aprendieran a reconocer individuos de su propia especie y contrarrestaran los efectos de ser criados por una especie distinta (p.e., en el emparejamiento con la especie que lo ha criado). Además, debido a que algunos críalos abandonan los nidos de Urraca después de que los adultos hayan abandonado la zona, estos podrían aprender la especie a la que pertenecen por los contactos con otros jóvenes en los grupos de críalos volantones. Estas ideas fueron testadas experimentalmente transportando pollos de Críalo desde Guadix a una zona, dentro del área de distribución del Críalo, pero con tasa de parasitismo muy baja (2,3%; Torres del Segre, Lérida), y a una zona en alopatría con el parásito (Freneusse, Paris), y en cada nido se metían 1 o 2 pollos (Soler y Soler, 1999). Todos los pollos experimentales procedían de nidos con un solo pollo de Críalo y, por tanto, no habían mantenido contacto con individuos de su misma especie antes del experimento. De acuerdo con la hipótesis de que las interacciones sociales entre adultos y jóvenes críalos son necesarias para que los jóvenes aprendieran la apariencia de los individuos de su especie (impronta), sólo los pollos experimentales llevados a Lérida, e introducidos de dos en dos en los nidos, formaron grupos de individuos procedentes de distintos nidos (dos grupos con 4 y 3 individuos) como ocurría en la zona de Guadix. Además, cuando estos dos grupos fueron detectados, también encontramos críalos adultos en los alrededores. Por otra parte, ninguno de los críalos experimentales que fueron puestos en nidos de Urraca sin la compañía de otro pollo de su especie, ni en Lérida ni en París, formaron grupos de jóvenes ni tampoco se detectaron individuos adultos en las proximidades de estos pollos (Soler y Soler, 1999). Estos resultados sugieren que los jóvenes de Críalo deben de aprender a reconocer los individuos de su propia especie. Es decir este reconocimiento no es innato en críalos. Además, los contactos entre adultos y jóvenes por un lado, y entre jóvenes procedentes de distintos nidos por otro, parecen esenciales para que este reconocimiento ocurra.
Referencias Arias de Reyna, L. (1998). Coevolution of great spotted cuckoo and its hosts. Pp. 129-142. En: Rothstein, S. I., Robinson, S. K. (eds.). Parasitic birds and their hosts, studies in coevolution. Oxford University Press, Oxford. Arias de Reyna, L., Recuerda, P., Corvillo, M., Aguilar, I. (1982). Reproducción del críalo (Clamator glandarius) en S. Morena Central. Doñana, Acta Vert., 9: 117-193. Arias de Reyna, L., Recuerda, P., Trujillo, J., Corvillo, M., Cruz, A. (1987). Territory in the great spotted cuckoo (Clamator glandarius). J. Ornithol., 128: 231-239. Bateson, P. (1979). How do sensitive periods arise and what are they for? Anim. Behav., 24: 470-486. Freeberg, T. M., King, A. P., West, M. J. (1995). Social malleability in cowbirds (Molothrus ater artemisiae): species and mate recognition in the first two years of life. J. Comp. Psychol., 109: 357-367. Gaston, A. J. (1976). Brood parasitism by the pied crested cuckoo (Clamator jacobinus). Journal of Animal Ecology, 45: 331-348. Kruijt, J. K., Cate, C. J. t., Meeuwissen, G. B. (1983). The influence of siblings on the development of sexual preferences of male zebra finches. Dev. Psychobiol., 16: 233-239. Lorenz, K. (1935). Der kumpan in der umwelt des vogels. J. Ornithol., 83: 137-213. Martínez, J. G., Burke, T., Dawson, D., Soler, J. J., Soler, M., Møller, A. P. (1998a). Microsatellite typing reveals mating patterns in the brood parasitic great spotted cuckoo (Clamator glandarius). Mol. Ecol., 7: 289-297. Martínez, J. G., Soler, J. J., Soler, M., Burke, T. (1998b). Spatial patterns of egg laying and multiple parasitism in a brood parasite: a non-territorial system in the great spotted cuckoo (Clamator glandarius). Oecologia, 117: 286-294. Mountfort, G., Ferguson-Lees, I. J. (1961). The Birds of the Coto Doñana. Ibis, 103: 86-109. Mundy, P. J., Cook, A. W. (1977). Observations on the breeding of the pied crow and great spotted cuckoo in Northern Nigeria. Ostrich, 48: 72-84. Soler, M., Palomino, J. J., Martínez, J. G., Soler, J. J. (1995c). Communal parental care by monogamous magpie hosts of fledgling great spotted cuckoos. Condor, 97: 804-810. Soler, M., Soler, J. J. (1991). Growth and development of great spotted cuckoos and their magpie host. Condor, 93: 49-54. Soler, M., Soler, J. J. (1999). Innate versus learned recognition of conspecifics in great spotted cuckoos Clamator glandarius. Anim. Cogn., 2: 97-102. Tinbergen, N. (1951). The study of instinct. Clarendon. Valverde, J. A. (1971). Notas sobre la biología reproductora del críalo (Clamator glandarius). Ardeola, (vol. Especial): 549-647.
Juan José Soler Fecha de publicación: 20-08-2003 Soler, J. J. (2016). Críalo Europeo - Clamator glandarius. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Morales, M. B. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|
|
|
|
|
|