|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Cabrera’s vole, description, size, variation.
Nombres comunes Los nombres comunes de la especie varían según el idioma y la región de origen. En castellano, además de topillo de Cabrera, la especie puede ser también denominada topillo Ibérico. Otros nombres comunes incluyen talpó de Cabrera (catalán); Cabrera lursagua (vasco); rato de Cabrera (portugués); campagnol de Cabrera, campagnol denté, campagnol méditerranéen, y campagnol iberique (francés); arvicola di Cabrera (italiano); Cabrera vole, Iberian vole, and Mediterranean vole (English); y Cabreramaus (alemán). Recientemente, el nombre común español Iberón (de Iberomys) ha sido propuesto en alternativa al topillo de Cabrera, principalmente enfocado hacia tareas de conservación (Cuenca-Bescós et al., 2014).
Origen y evolución El topillo de Cabrera evolucionó a partir de la radiación de los primeros topillos con dientes de raíces abiertas, dentro del grupo Allophaiomys (Chaline 1970), que estaba ampliamente distribuido en el sur de Europa durante el inicio del Pleistoceno (Agusti 1991), y cuyo ancestro habría sido, presumiblemente, una rata de agua del Plioceno, Mimomys (Chaline y Graf 1988; Ruiz-Bustos y Sesé 1985). El linaje anagenético Iberomys, al cual pertenece el topillo de Cabrera (Chaline 1972a), se distingue de otros Arvicolidae con dientes de raíces abiertas, por sus molares marcadamente asimétricos y en forma de triángulo, en particular el m1 (Cabrera-Millet et al., 1983; Cuenca-Bescós y Laplana 1995). La especie M. huescarensis (Ruiz Bustos, 1988) es la primera representante del linaje Iberomys, apareciendo en la Península Ibérica a finales del Pleistoceno. Durante el Pleistoceno medio, M. huescarensis fue reemplazado por M. brecciensis (Giebel 1847), que se convirtió en el topillo más abundante y ampliamente distribuido en toda la región Iberoccitana (Península Ibérica y la región mediterránea del sur de Francia). M. brecciensis (sinónimo de Hipudaeus brecciensis, Giebel 1847; Forsyth Major 1905) es el antepasado directo del topillo de Cabrera (Cabrera-Millet et al 1983), que poco a poco redujo su rango de distribución desde finales del Pleistoceno hasta la actualidad (López- Martínez, 2009; Pita et al., 2014). Las diferencias en la morfología de las mandíbulas y dentición entre el topillo de Cabrera y M. brecciensis fueron descritas por Ayarzaguena y López-Martínez (1976) y por Sesé et al. (2011b), sugiriendo que el topillo de Cabrera es de mayor tamaño que M. brecciensis. Análisis filogenéticos basados en secuencias del gen citocromo-b son consistentes con la idea de que, entre los Arvicolinae, el linaje de Iberomys parece ser muy antiguo (Jaarola et al., 2004).
Taxonomía y nomenclatura El topillo de Cabrera fue descrito en 1906 por Oldfield Thomas (Museo de Historia Natural de Londres) basándose en un solo espécimen recogido el mismo año por Manuel Martínez de la Escalera en la Sierra de Guadarrama (Thomas, 1906). El nombre “cabrerae” fue dedicado en honor a Ángel Cabrera Latorre (1879-1960), mastozoólogo del Museo Nacional de Ciencias Naturales en Madrid, y que fue discípulo de Thomas (Tellado y Molina, 2010). Taxonómicamente, el topillo de Cabrera ha sido incluido en diferentes subgéneros a lo largo del tiempo (Pita et al., 2014). En 1987, Pavlinov y Rossolimo lo incluyeron dentro del subgénero Microtus (Musser y Carleton, 2005). Sin embargo, basado en caracteres morfológicos arcaicos, Chaline (1972a) utilizó el subgénero Iberomys para la especie, aunque la incluyese más tarde en el subgénero Arvalomys, junto con M. arvalis, M. agrestis y otros topillos americanos (Chaline, 1974). El topillo de Cabrera fue también incluido en el subgénero Agricola por Zagorodnuyuk en 1990 (Musser y Carleton, 2005), aunque los datos filogenéticos recientes (Jaarola et al., 2004) soportan su reclasificación bajo el subgénero Iberomys, como propuso anteriormente Chaline (1972). El topillo de Cabrera es el único representante existente del subgénero Iberomys. Otras especies incluidas en este subgénero son M. huescarensis (Mazo, 1985) y M. brecciensis (Giebel, 1947), antepasados del topillo de Cabrera (Cabrera-Millet et al., 1983). El subgénero Iberomys ha sido considerado como un género distinto de Microtus basándose en la morfología de m1 (Cuenca-Bescós y Laplana, 1995, Laplana y Cuenca-Bescós, 1998), y algunos autores se refieren al topillo de Cabrera como Iberomys cabrerae (López-García, 2008). La propuesta de elevar el subgénero Iberomys al nivel de género se ha reforzado recientemente (Cuenca-Bescós et al., 2014), fundamentada en sus caracteres morfológicos distintivos de otros Microtus, su historia paleogeográfica, y ciertas características citogenéticas y ecológicas. Todavía, faltan dados genéticos que soporten el cambio del subgénero Iberomys al nivel de género (Jaarola et al., 2004). El topillo de Cabrera ha sido considerado como una especie distinta de M. dentatus (Miller, 1910; Cabrera, 1914), aunque también ha sido reconocido como una subespecie de M. cabrerae (Ellerman y Morrison-Scott, 1951, Gromov y Pavlínov, 1992), con M. c. cabrerae ocurriendo principalmente en el norte y centro de la Península Ibérica, y M. c. dentatus predominando en el sureste. Sin embargo, en base a estudios de morfología (Niethammer et al., 1964; Ayarzaguena y López-Martínez 1976), electroforesis (Cabrera-Millet et al., 1982), y biometría (Ventura et al., 1998), esta variación subespecífica no se acepta en la actualidad, y se considera a M. dentatus como un sinónimo de M. cabrerae. El topillo de Cabrera ha sido también referido como M. cabrerai por Trouessart (1910), aunque la forma correcta de escribir el nombre de la especie es la latinización original de Thomas, de Cabrera para cabrerae.
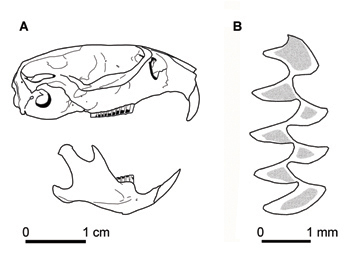
Diagnosis Las especies de Microtus existentes actualmente en la Península Ibérica incluyen: el topillo lusitano M. lusitanicus (al norte de Iberia), el topillo mediterráneo M. duodecimcostatus (al sur de Iberia), el topillo pirenaico M. gerbei (en los Pirineos y montañas del este de Cantabria), el topillo agreste M. agrestis (al norte de Iberia), y el topillo campesino M. arvalis (al norte y centro de Iberia). De forma similar a estas especies, la longitud de la cola del topillo de Cabrera es aproximadamente un tercio de la longitud del cuerpo (Niethammer et al 1964; Madureira y Ramalhinho 1981). Sin embargo, el topillo de Cabrera puede distinguirse fácilmente de todas estas especies por su mayor tamaño (Pita et al., 2014). El rango de dimensiones físicas de topillos del género Microtus en la Península Ibérica referidas en Fernández-Salvador (2002, 2007b) indican que la masa corporal media del topillo de Cabrera es de aproximadamente 1,3 a 4,9 veces mayor que las del topillo lusitano, topillo mediterráneo y topillo pirenaico; y alrededor de 1,0 a 3,8 mayor que las del topillo agreste y topillo campesino. La longitud del cuerpo y la cola son, respectivamente, de alrededor de 1,1-1,7 y 0,9-2,7 veces más largas que las del topillo lusitano, topillo mediterráneo y topillo pirenaico; y alrededor de 0,8-1,9 y 0,7-1,8 veces más que las del topillo agreste y topillo campesino. Las orejas son relativamente largas, con una longitud de aproximadamente 1,3-2,5 veces más larga que en el caso de topillo lusitano, topillo mediterráneo y topillo pirenaico; y alrededor de 0,9-1,6 veces más larga que las del topillo agreste y topillo campesino. También la longitud del pie es mayor en el topillo de Cabrera, siendo una característica discriminante particularmente útil (en general >20 mm para el topillo de Cabrera y <20 mm para todas las demás especies Ibéricas de Microtus). Al igual que en el topillo campesino, el topillo de Cabrera tiene 6 almohadillas plantares, mientras que las demás especies Ibéricas de Microtus tienen 5. El topillo de Cabrera también exhibe características craneales diferenciales (Fig. 1A). Su mayor tamaño, una cápsula incisiva más pequeña en la mandíbula, y un complejo tercer molar (con 3 y 4 prismas en m3 and M3, respectivamente) ayudan a distinguirlo de otras especies de Microtus Ibéricos (Niethammer et al., 1964; Engels, 1972). Sin embargo, la principal característica que distingue al topillo de Cabrera y el linaje Iberomys de otros Arvicolinae con dientes de raíces abiertas, es la marcada asimetría transversal de los molares, en particular el m1 (cerca de 1,20-1,43 largo), con triángulos linguales más anchos que los triángulos labiales (Fig. 1B). Además, los ángulos salientes son más agudos, contrastando con los de otras especies de Microtus, Chionomys y Arvicola, que presentan ángulos salientes redondeados (Niethammer et al. 1964; Ayarzaguena and Lopéz-Martinez 1976; Cuenca-Bescós and Laplana 1995; Cuenca-Bescós et al. 2014).
Figura 1. (A) Cráneo y mandíbula; y (B) primero molar inferior derecho (m1) de topillo de Cabrera (C) R. Pita
Las características esqueléticas post-craneales con respecto a otros Microtus ibéricos incluyen un húmero más largo y más robusto, con cabeza articular más pequeña y menos sobresaliente; un proceso del olécranon del cúbito más largo y más robusto; y la pelvis más robusta, con un ilion más largo y más ancho (Cuenca-Bescós et al. 2014).
Descripción El topillo de Cabrera es una especie de arvicolino de tamaño mediano, aunque entre los miembros de su género posee una de las masas corporales más grandes, con una masa media de alrededor de 52 g, y un rango de entre 30-78 g (Ventura et al., 1998; Figura 2). El pelo es largo y grueso, de color oliváceo-pardusco dorsalmente y amarillento ventralmente (Thomas, 1906; Engels, 1942; Niethammer et al., 1964). Como en otros arvicolinos, los juveniles de topillo de Cabrera son de un color más oscuro que los adultos y sub-adultos. Las orejas son pequeñas y casi completamente cubiertas de pelo, y la cola es corta y ligeramente bicolor (blanco-pardusco dorsalmente, blanco ventralmente). Según Niethammer (1982), aunque basado en un pequeño tamaño de muestra, el pelaje es más denso y la longitud del pelo y de las vibrisas es mayor en M. cabrerae que en M. arvalis y que en M. agrestis. Las patas posteriores son largas y grisáceas (Thomas 1906; Engels 1942). No hay diferencias físicas evidentes entre machos y hembras, excepto en las dimensiones de la pelvis en individuos adultos (Ayarzaguena y Cabrera 1976). La fórmula dental es 2(I 1/1, C 0/0, P 0/0, M 3/3), total 16 (Pita et al., 2014).
Biometría La variabilidad morfométrica fue descrita en detalle por Ventura et al. (1998), a partir de 148 especímenes recogidos entre 1977 y 1981 en varias localidades de diferentes regiones geográficas de España (provincias de Albacete, Cáceres, Cuenca, Madrid, Jaén, Zaragoza y Huesca; Tabla 1), no existiendo diferencias en ninguna de las mediciones morfológicas y craneales entre sexos o entre localidades.
Tabla 1. Medidas corporales y del cráneo del topillo de Cabrera (en mm) de acuerdo con Ventura et al. 1998. a individuos de las provincias de Albacete, Cáceres, Cuenca, Madrid, y Jaén; b individuos de las provincias de Albacete, Cáceres, Cuenca, Madrid, Jaén, Zaragoza, y Huesca; c individuos de las provincias de Albacete, Cáceres, Madrid, Jaén, Zaragoza, y Huesca.
Las relaciones entre las mediciones craneales y la masa corporal revelan un escalonamiento isométrico con la longitud corporal, una alometría positiva para la longitud de la cola, y una alometría negativa para las longitudes condilobasal, de diastema superior y mandíbula (Ventura et al., 1998). Varias medidas craneales presentan escalonamiento isométrico con la longitud de diastema superior, como, por ejemplo, las alturas articulares y interparietal-foramen magnum, las anchuras rostral y cigomática, y las longitudes del foramen condilobasal, nasal, incisivo y condiloincisivo (Ventura et al., 1998). Es también aparente una alometría negativa entre la longitud del diastema superior y los bordes dentales maxilares y mandibulares, la altura del cráneo, las longitudes occipitonasales y mandibulares y las anchuras de la caja craneana, los nasales y occipitales (Ventura et al., 1998). La relación entre la anchura del cigomático y la longitud del diastema indica que la ampliación de los arcos cigomáticos es paralela a la elongación del viscerocráneo (Ventura et al., 1998). El crecimiento relativamente importante de un complejo arco cigomático, está probablemente asociado con el fuerte desarrollo del masetero lateral durante la vida postnatal. De forma similar a otras especies de roedores, el pronunciado desarrollo de los músculos pterigoideos maseteros y laterales está relacionado con un desarrollo predominante del proceso articular sobre el crecimiento de la mandíbula (Ventura et al., 1998).
Cariotipo Los análisis citogenéticos revelan que el topillo de Cabrera posee un número diploide de 54 cromosomas, y un número fundamental de 64 (Díaz de la Guardia et al., 1979; Palacios y Cabrera 1979; Burgos et al., 1988a). Entre las especies de Microtus ibéricas contemporáneas sólo M. gerbei tiene el mismo número diploide (Musser y Carleton 1993). El cariotipo del topillo de Cabrera fue originalmente descrito por Díaz de la Guardia et al. (1979), el cual está constituido por 3 pares de cromosomas submetacéntricos grandes (pares 1, 2 y 3), 1 par de pequeños cromosomas metacéntricos (par 4), y 22 pares de cromosomas acrocéntricos (pares 5 A 26) que muestran diferencias graduales de tamaño. Sin embargo, Palacios y Cabrera (1979) demostraron que puede haber variabilidad en la morfología cromosómica. Las técnicas de bandeo cromosómico han mostrado que los autosomas del topillo de Cabrera tienen unas bandas-C centrómeras más grandes que las de otras especies ibéricas de arvicolinos (Burgos et al., 1988a; Fernández et al., 2001). Sin embargo, una de las características más importantes del cariotipo del topillo de Cabrera es la presencia de cromosomas sexuales “gigantes”, con grandes secciones de heterocromatina constitutiva (Díaz de la Guardia et al., 1979; Burgos et al., 1988a; Marchal et al., 2004); lo cual, entre los Microtus ibéricos, apenas puede encontrarse en M. agrestis (2n = 50, Burgos et al., 1988a; Jiménez et al., 1991; Marchal et al., 2004; Giménez et al., 2012). La estructura de heterocromatina en los cromosomas sexuales del topillo de Cabrera es altamente heterogénea, muy probablemente como resultado de una rápida amplificación de bloques de secuencias repetitivas dentro de las diversas secciones, cada una de origen y composición diferentes (Burgos et al 1988c; Bullejos et al., 1996; Fernández et al., 2001; Marchal et al., 2004). Se han descrito 6 subtipos diferentes de heterocromatina en los cromosomas sexuales del topillo de Cabrera, 4 en el cromosoma X, y 2 en el cromosoma Y (Burgos et al., 1988c; Fernández et al., 2001; Marchal et al., 2004). Otra característica de la citogenética molecular del topillo de Cabrera es la presencia de múltiples copias del gen SRY, en lugar de la única copia observada en la mayoría de las especies de mamíferos (Bullejos et al., 1997). Aunque en varias especies de Microtus se encuentran múltiples copias del gen SRY (Bullejos et al., 1999), en el topillo de cabrera el gen SRY puede estar presente tanto en machos como en hembras (Fernández et al., 2002), y no sólo en la región no recombinante del Y (Acosta et al., 2010). Debido a que el cromosoma Y está normalmente presente en los machos y ausente en las hembras, y que algunos casos de reversión de sexo han sido descritos en hembras XY (Burgos et al., 1988b), es posible que una copia funcional del gen SRY localizada en el cromosoma Y pueda actuar como el gen determinante de la formación de los testículos (Fernández et al., 2002). Por otro lado, las copias SRY de los cromosomas X deben haber estado siempre inactivas, ya que la presencia de copias funcionales de este gen en las hembras daría origen a machos XX estériles y, por consiguiente, se habrían perdido los cromosomas X con copias SRY de la población (Fernández et al., 2002).
Variación geográfica La masa corporal y las biometrías craneales y post-craneales descritas en Ventura et al. (1998) no evidencian relaciones geográficas significativas. Además, a pesar de la gran variabilidad en los triángulos T1 y T2 del m3 (que pueden ser confluentes, semi-confluentes, o aislados), estos morfotipos no están tampoco relacionados con populaciones geográficamente distintas (Ventura et al. 1998). Sin embargo, la variación de las secuencias en el ADN mitocondrial reveló la existencia de dos linajes principales bien diferenciados, denominados este y oeste (Barbosa et al., 2017). El linaje estepresenta una mayor diversidad nucleotídica, pero menos haplotipos, los cuales están particularmente restringidos geográficamente. Esta diferenciación este-oeste no parece coincidir con las variaciones en el ADN nuclear, las cuales indicaron menor diversidad nucleotídica en el grupo pre-pirenaico (Barbosa et al., 2017).
Registro fósil Los lugares arqueológicos y paleontológicos con fósiles de Iberomys (heuscarensis-brecciensis-cabrerae) han sido inventariados exhaustivamente en Garrido-García y Soriguer-Escofet (2012) y Laplana y Sevilla (2013). Los fósiles de M. huescarensis se han encontrado en 4 sitios de España: cerca de Atapuerca (Burgos, Laplana y Cuenca-Bescós, 1998; Cuenca-Bescós et al., 1999), Vallparadis, Terrassa (Barcelona, Martínez et al., 2010), Almenara-Casablanca (Castellón, Agusti et al., 2011) y Caravaca de la Cruz (Murcia, Walker et al., 2013). Los fósiles de M. brecciensis se han encontrado en más de 50 sitios de la Península Ibérica (incluyendo Portugal, España y Gran Bretaña-Gibraltar), Francia, e Italia (Garrido-García y Soriguer-Escofet, 2012). Además, fósiles descritos como Hipudaeus brecciensis fueron encontrados en la Península Balcánica (Marjan, Croacia, Vuletic, 1953), aunque no se dispone de información sobre las medidas craneales o del tamaño y estructura de los molares para su confirmación. Los subfósiles de topillo de Cabrera son relativamente comunes en muchos depósitos del Pleistoceno-Holoceno tardío en la Península Ibérica y sur de Francia. Al contrario de su antepasado M. brecciensis, el topillo de Cabrera está ausente de Italia y de la región cantábrica de España (Sesé et al., 2011a). Los restos fósiles del Pleistoceno tardío y Holoceno encontrados en el sur de Francia apoyan la contracción general de la parte oriental de su rango de distribución hasta el presente (Pita et al., 2014).
Indicios Los nidos de topillo de Cabrera están normalmente ocultos en masas de vegetación verde, normalmente en la base de juncos o arbustos, aunque si la vegetación no ofrece suficiente protección pueden situarse al final de una galería subterránea (Garrido-García y Soriguer, 2014, Figura 2). Las huellas raramente se marcan en el suelo compactado de los caminos típicamente usados por el topillo de Cabrera (Garrido-García y Soriguer, 2014).
Figura 2 . Nido de topillo de Cabrera con crías. Vila Nova de Milfontes, Portugal. (C) R. Pita
Figura 3. Excrementos/letrina de topillo de Cabrera (C) D. Peralta
Los excrementos (con cerca de 4-8 mm de largo) depositados en letrinas (Figura 3), y los restos de comida (hierba cortada) que aparecen a lo largo de los caminos de tránsito hechos en la hierba, constituyen los indicios que se encuentran con mayor frecuencia, siendo comúnmente usados para muestrear poblaciones locales de topillo de Cabrera (Pita et al., 2014). Los caminos son más estrechos que los de la rata de agua (Arvicola sapidus) y más largos que los del topillo mediterráneo. Los excrementos son cilíndricos, con los extremos redondeados, y típicamente compuestos de materia vegetal, presentando un color verdoso o pardusco (dependiendo de la alimentación y tiempo de decaimiento). Los excrementos de A. sapidus, que también forma letrinas, son más grandes, especialmente en anchura (Garrido-García y Soriguer, 2014). En zonas de simpatría con el topillo agreste, podrían ser necesarios procedimientos genéticos para diferenciar los excrementos del topillo de Cabrera. De hecho, el reciente desarrollo de marcadores genéticos específicos para la especie, así como los recientes avances en protocolos de extracción de ADN basados en el muestreo genético no invasivo de excrementos, destacan como buenas alternativas para ampliar el repertorio de técnicas de muestreo del topillo de Cabrera (Alasaad et al, 2010, 2011, 2012, Barbosa et al., 2013), ya que la captura de animales vivos es cara logísticamente, y normalmente resulta insuficiente para estimar parámetros demográficos (Landete-Castilejos et al., 2000; Fernández-Salvador et al., 2005).
Referencias Acosta, M. J., Marchal, J. A., Romero-Fernández, I., Megías-Nogales, B., Modi, W. S., Sánchez, A. (2010). Sequence analysis and mapping of the Sry gene in species of the subfamily Arvicolinae (Rodentia). Sexual Development, 4: 336-347. Agustí, J. (1991). The Allophaiomys complex in southern Europe. Geobios, 25: 133-144. Agustí, J., Santos-Cubedo, A., Furió, M., De Marfa, R., Blain H.-A., Oms, O., Sevilla, P. (2011). The late Neogene-early Quaternary small vertebrate succession from the Almenara-Casablanca karst complex (Castellón, Eastern Spain): Chronologic and paleoclimatic context. Quaternary International, 243: 183-191. Alasaad, S., Soriguer, R.C., Jowers, M. J., Marchal, J. A., Romero, I., Sánchez, A. (2010). Applicability of mitochondrial DNA for the identification of Arvicolid species from faecal samples: a case study from the threatened Cabrera's vole. Molecular Ecology Resources, 11: 409-414. Alasaad, S., Sánchez, A., Marchal, J. A., Píriz, A., Garrido-García, J. A., Carro, F., Romero, I., Soriguer, R. C., 2011). Efficient identification of Microtus cabrerae excrements using noninvasive molecular analysis. Conservation Genetics Resources, 3: 127-129. Alasaad, S., A. Sánchez, A., García-Mudarra, J. L., Pérez, J. M., Marchal, J. A., Romero, I., Garrido-García, J. A., Soriguer, R. C. (2012). Single-tube HotSHOT technique for the collection, preservation and PCR-ready DNA preparation of fecal samples: the threatened Cabrera's vole as a model. European Journal of Wildlife Research, 58: 345-350. Ayarzaguena, J., Cabrera, M. (1976). Dimorfismo sexual en Microtus cabrerae en base a los caracteres de su pelvis. Doñana, Acta Vertebrata, 3: 107-108. Ayarzaguena, J., López-Martínez, N. (1976). Estudio filogenético y comparativo de Microtus cabrerae y Microtus brecciensis. Doñana, Acta Vertebrata, 3: 181-204. Barbosa, S., Paupério, J., Herman, J. S., Ferreira, C.M., Pita, R., Vale-Gonçalves, H., Cabral, J.A., Garrido-García, J.A., Soriger, R. C., Beja, P., Mira, A., Alves, P. C., Searle, J. B. (2017). Endemic species may have complex histories: within-refugium phylogeography of an endangered Iberian vole. Molecular Ecology, 26 (3): 951-967. Bullejos, M., Burgos, M., Jiménez, R., Sánchez, A., Díaz de la Guardia, R. (1996). Distribution of sister chromatid exchanges in different types of chromatin in the X chromosome of Microtus cabrerae. Experientia, 52: 511-515. Bullejos, M., Sánchez, A., Burgos, M., Hera, C., Jiménez, R., Díaz de la Guardia, R. (1997). Multiple, polymorphic copies of SRY in both males and females of the vole Microtus cabrerae. Cytogenetics and Cell Genetics, 79: 167-171. Bullejos, M., Sánchez, A., Burgos, M., Jiménez, R., Díaz de la Guardia, R. (1999). Multiple mono- and polymorphic Y-linked copies of the SRY HMG-box in Microtidae. Cytogenetics and Cell Genetics, 86: 46-50. Burgos, M., Jiménez, R., Díaz de la Guarda, R. (1988a). Comparative study of G- and C-banded chromosomes of five species of Microtidae: a chromosomal evolution analysis. Genome, 30: 540-546. Burgos, M., Jiménez, R., Díaz de la Guardia, R. (1988b). XY females in Microtus cabrerae (Rodentia, Microtidae): a case of possibly Y-linked sex reversal. Cytogenetics and Cell Genetics, 49: 275-277. Burgos, M., Jiménez, R., Olmos, D. M., Díaz de la Guardia, R. (1988c). Heterogeneous heterochromatin and size variation in the sex chromosomes of Microtus cabrerae. Cytogenetics and Cell Genetics, 47: 75-79. Cabrera, A. (1914). Fauna Ibérica: Mamíferos. Museo Nacional de Ciencias Naturales, Madrid. Cabrera-Millet, M., Britton-Davidian, J., Orsini, P. (1982). Génétique biochimique comparée de Microtus cabrerae Thomas, 1906 et de trois autres espèces d’Arvicolidae mediterranéens. Mammalia, 46: 381-388. Cabrera-Millet, M., López-Martínez, N., Michaux, J. (1983). Un exemple de lignee endemique iberoccitane, les campagnols Microtus brecciensis et Microtus cabrerae (Mammalia, Rodentia): Etude phylogenetique et contexte ecologique d’un phenomene evolutif recent. Pp 69-83. En: (Buffetaut, E., Mazin, J. M., Salmon, E. (Eds.). Actes du Symposium Paleontologique. Montbélliard, France. Chaline, J. (1970). La signification des Rongeurs dans les dépôts quaternaires. Bulletin de l'Association Française pour l'Étude du Quaternaire, 7: 229-241. Chaline, J. (1972). Les Rongeurs du Pleistocene moyen et superieur de France. Systematique, Biostratigraphie, Paleoclimatologie. Cahiers de Paleontologie. C.N.R.S. París, France. Chaline, J. (1974). Esquisse de l’évolution morphologique, biométrique et chromosomique du genre Microtus (Arvicolinae, Rodentia) dans le Pléistocène de l’hémisphère nord. Bulletin de la Société de Géologie France, sér. 7, 14: 440-450. Chaline, J., Graf, J. D. (1988). Phylogeny of the Arvicolidae (Rodentia): Biochemical and paleontological evidence. Journal of Mammalogy, 69: 22-33. Cuenca-Bescós, G., Laplana, C. (1995). Evolución de Iberomys (Arvicolidae, Rodentia, Mammalia) durante el Cuaternario español. Pp. 69–72. En: López, G., Obrador, A., Vicens, E. (Eds.). XI Jornadas de Paleontología. Universidad Autónoma de Barcelona, Barcelona. Cuenca-Bescós, G., López-García, J. M., Galindo-Pellicena, M. A., García-Perea, R., Gisbert, J., Rofes, J., Ventura, J. (2014). The Pleistocene history of Iberomys an endangered rodent from south Western Europe. Integrative Zoology, 9 (4): 481-497. Cuenca-Bescós, G., Laplana, C., Canudo, J. I. (1999). Biochronological implications of the Arvicolidae (Rodentia, Mammalia) from the Lower Pleistocene hominid-bearing level of Trinchera Dolina 6 (TD6, Atapuerca, Spain). Journal of Human Evolution,37: 353-373. Díaz de la Guardia, R., Pascual, L., Orozco, J. V. (1979). The karyotype of Microtus cabrerae Thomas, another species with giant sex chromosomes. Experientia, 35: 741-742. Ellerman, J. R., Morrison-Scott, T. C. S. (1951). Checklist of Paleartic and Indian Mammals 1758 to 1946. British Museum (Natural History), London. Engels, H. (1972). Kleinsäuger aus Portugal. Bonner Zoologische Beitrage, 21: 89-118. Fernández, R., Barragán, M. J., Bullejos, M., Marchal, J. A., Martínez, S., Díaz de la Guardia, R., Sánchez, A. (2001). Molecular and cytogenetic characterization of highly repeated DNA sequences in the vole Microtus cabrerae. Heredity, 87: 637-646. Fernández, R., Barragán, M. J. L., Marchal, J. A., Martínez, S., Díaz la Guardia, R., Sánchez, A. (2002). Mapping the SRY gene in Microtus cabrerae: A vole species with multiple SRY copies in males and females. Genome, 45: 600-603. Fernández-Salvador, R. (2002). Microtus cabrerae Thomas, 1906. Topillo de Cabrera. Pp. 386-389. En: Palomo, L. J., Gisbert, J. (Eds.). Atlas de los mamíferos terrestres de España. Dirección General de Conservación de la Naturaleza – Ministerio de Medio Ambiente, Sociedad Española para la Conservación y Estudio de los Mamíferos, Sociedad Española para la Conservación y Estudio de los Murciélagos, Madrid. 564 pp. Fernández-Salvador, R. (2007b). Microtus cabrerae Thomas, 1906. Topillo de Cabrera. Pp. 429-433. En: Palomo, J. L., Gisbert, J., Blanco, J. C. (Eds.). Atlas y Libro Rojo de los mamíferos terrestres de España. Dirección General para la Biodiversidad, SECEM-SECEMU, Madrid. 586 pp. Fernández-Salvador, R., Ventura, J., García-Perea, R. (2005). Breeding patterns and demography of a population of the Cabrera vole, Microtus cabrerae. Animal Biology, 55: 147-161. Forsyth Major, C. I. (1905). Rodents from the Pleistocene of the Western Mediterranean Region. Geological Magazine, 5: 462-467.Garrido-García, J. A., Soriguer-Escofet, R. C. (2012). Cabrera's Vole Microtus cabrerae Thomas, 1906 and the subgenus Iberomys during the Quaternary: evolutionary implications and conservation. Geobios, 45: 437-444.Garrido-García, J. A., Soriguer, R. C. (2014) Topillo de Cabrera Iberomys cabrerae (Thomas, 1906) En: Calzada. J., Clavero, M., Fernández, A. (Eds.). Guía virtual de los indicios de los mamíferos de la Península Ibérica, Islas Baleares y Canarias. Sociedad Española para la Conservación y Estudio de los Mamíferos (SECEM). http://www.secem.es/guiadeindiciosmamiferos/Giebel, C. G. (1847). Fauna der Vorwelt mit steter Berücksichtigung der lebenden Thiere. Erster Band: Wirbelthiere, Erster Abtheilung: Säugethiere. F. A. Brockhaus, Luipzig.Giménez, M., Paupério, J., Alves, P.C., Searle J. B. (2012). Giant sex chromosomes retained within the Portuguese lineage of the field vole (Microtus agrestis). Acta Theriologica, 57: 377-382Gromov, I. M., Polyakov, I. Y. (1992). Fauna of the USSR, mammals, voles (Microtinae), Vol. III, No. 8. English translation; originally published in Russian, 1977, Nauka: Moskow-Leningrado. D. Siegel-Causey and R. S. Hoffman (Eds.). Smithsonian Institute Libraries, Washington, D. C. Jaarola, M., Martínková, N., Gündüz, I., Brunhoff, C., Zima, J., Nadachowski, A., Amori, G., Bulatova, N. S., Chondropoulos, B., Fraguedakis-Tsolis, S., González-Esteban, J., López-Fuster, M. J., Kandaurov, A. S., Kefelioğlu, H., Mathias, M. L., Villate, I., Searle, J. B. (2004). Molecular phylogeny of the speciose vole genus Microtus (Arvicolinae, Rodentia) inferred from mitochondrial DNA sequences. Molecular Phylogenetics and Evolution, 33: 647-663. Jiménez, R., Carbero, M., Burgos, M., Sánchez, A., Díaz de la Guardia, R. (1991). Achiasmatic giant sex chromosomes in the vole Microtus cabrerae (Rodentia, Microtidae). Cytogenetics and Cell Genetics, 57: 56-58. Landete-Castillejos, T., Andrés-Abellán, M., Argandoña, J. J., Garde, J. (2000). Distribution of the Cabrera vole in its first reported areas reassessed by life-trapping. Biological Conservation, 94: 127-130.Laplana, C., Cuenca-Bescós, G. (1998). Presisiones sobre la systemática de Iberomys Chaline, 1972 (Arvicolidae, Rodentia, Mammalia) del Pleistoceno inferior de la Sierra de Atapuerca (Burgos, España). Geogaceta, 24: 183-186.Laplana, C., Sevilla, P. (2013). Documenting the biogeographic history of Microtus cabrerae through its fossil record. Mammal Review, 43 (4): 309-322. López-García, J. M. (2008). Evolución de la diversidad taxonómica de los micromamíferos en la Península Ibérica y cambios Paleoambientales durante el Pleistoceno Superior. Tesis doctoral. Universitat Rovira i Virgili, Tarragona. López-Martínez, N. (2009). Time asymmetry in the palaeobiogeographic history of species. Bulletin de la Société Géologique de France, 180: 45-55. Madureira, M. L., Ramalhinho, M. G. (1981). Notas sobre a distribuição, diagnose e ecologia dos insectivora e rodentia portugueses. Arquivos do Museu Bocage, Série A, 1: 165-263. Martínez, K., Garcia, J., Carbonell, E., Agustí, J., Bahain, J. J., Blain, H. A., Burjachs, F., Cáceres, I., Duval, M., Falguères, C., Gómez, M., Huguet, R. (2010). A new Lower Pleistocene archeological site in Europe (Vallparadís, Barcelona, Spain). Proceedings of the National Academy of Sciences, 107: 5762-5767. Mazo, A. V., Sesé, C., Ruiz Bustos, A., Peña, J. A. (1985). Geología y peleontología de los yacimientos Plio-Pleistocenos de Huéscar (Depresión de Guadix-Baza, Granada). Estudios Geológicos, 41: 467-493. Marchal, J. A., Acosta, M. J., Nietzel, H., Sperling, K., Bullejos, M., Díaz de la Guardia, R., Sánchez, A. (2004). X chromosome painting in Microtus: Origin and evolution of the giant sex chromosomes. Chromosome Research, 12: 767-776. Miller, G. S. (1910). Description of six new European mammals. Annals of the Magazine of Natural History, Series 10, 6: 458-461. Musser, G. G., Carleton, M. D. (2005). Superfamily Muroidea. Pp. 894–1531. En: Wilson, D. E., Reeder, D. M. (Eds.). Mammal species of the world: a taxonomic and geographic reference. 3rd ed. Johns Hopkins University Press, Baltimore. Niethammer, J. (1982). Microtus cabrerae Thomas, 1906 – Feldmaus. Pp. 340-348. En: Niethammer, J., Krapp, F. (Eds.). Handbuch der Säugetiere Europas. Band 2/I. Rodentia II (Cricetidae, Arvicolidae, Zapodidae, Spalacidae, Hystricidae, Capromyidae), Akademische Verlagsgesellschaft, Wiesbaden. 649 pp. Niethammer, J., Niethammer, G., Abs, M. (1964). Ein Beitrag zur Kenntnis der Cabreramaus (Microtus cabrerae Thomas, 1906). Bonner Zoologische Beiträge, 3/4: 127-148. Palacios, F., Cabrera, M. (1979). Descripción de los cromosomas del toppillo Ibérico (Microtus cabrerae). Doñana, Acta Vertebrata, 6: 132-136. Pita, R., Mira, A., Beja, P. (2014). Microtus cabrerae (Rodentia: Cricetidae). Mammalian Species, 46 (912): 48-70. Ruiz Bustos, A. (1988). Estudio sobre los Arvicólidos Cuaternarios. Paleomammalia, 2: 1-89. Ruiz Bustos, A., Sesé, C. (1985). Evolución de los generos Minomys, Arvicola y Allophaiomys (Arvicolidae, Rodentia, Mammalia), en el Plioceno y Pleistoceno de la Peninsula Ibérica. Estudios Geológicos, 41: 99-104. Sesé, C., Panera, J., Rubio-Jara, S., Pérez-González, A. (2011a). Micromamíferos del Pleistoceno Medio y Pleistoceno Superior en el Valle del Jarama: Yacimientos de Valdocarros y HAT (Madrid). Estudios Geológicos, 67: 131-151. Sesé, C., Rubio-Jara, S., Panera, J., Pérez-González, A. (2011b). Micromamíferos del Pleistoceno Superior del yacimiento de PRERESA en el valle del Manzanares y su contribución a la reconstrucción paleoambiental de la cuenca de Madrid durante el Pleistoceno. Estudios Geológicos, 67: 471-494. Tellado, J. M., Molina, J. (2010). “Un mes en Londres”: Angel Cabrera Latorre at the British Museum (Natural History) and the launch of an international career. Archives of Natural History, 37: 1-18. Thomas, O. (1906). A new vole from Spain. Annals of the Magazine of Natural History, Series 7 17: 576-577. Trouessart, E. L. (1910). Conspectus mammalium Europae. Faune des mammifères d’Europe. Friedländer & Sohn, Berlin. Ventura, J., López-Fuster, M. J., Cabrera-Millet, M. (1998). The Cabrera vole, Microtus cabrerae, in Spain: A biological and morphometric approach. Netherlands Journal of Zoology, 48: 83-100. Vuletic, A. (1953). Brèche diluviale osseuse à Marjan. Biljeske Notes, 7: 1-3. Walker, M. J., López-Martínez, M. V., Carrión-García, J. S., Rodríguez-Estrella, T., San-Nicolás-del-Toro, M., Schwenninger, J. L., López-Jiménez, A., Ortega-Rodrigáñez, J., Haber-Uriarte, M., Polo-Camacho, J. L., García-Torres, J., Campillo-Boj, M., Avilés-Fernández, A., Zack, W. (2013). Cueva Negra del Estrecho del Río Quípar (Murcia, Spain): A late Early Pleistocene hominin site with an “Acheulo-Levalloiso-Mousteroid” Palaeolithic assemblage. Quaternary International, 294: 135-159.
Ricardo Pita1, Juan José Luque-Larena2, Pedro Beja3, António Mira1 Fecha de publicación: 26-09-2017 Pita, R., Luque-Larena, J. J., Beja, P., Mira, A. (2017). Topillo de Cabrera – Microtus cabrerae. En: Enciclopedia Virtual de los Vertebrados Españoles. Sanz, J. L., Barja. I. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||