|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Storm Petrel, description, size, variation, moult.
Identificación y morfología El paíño europeo (Hydrobates pelagicus, Figura 1) es, sin lugar a dudas, una de las aves marinas más esquivas debido a sus hábitos nocturnos y pelágicos. Descrita por Linnaeus en 1758 como Procellaria pelagica, años después, en 1822, Friedich Boie le cambió el género a Hydrobates. El nombre científico del paíño viene del griego y está relacionado con su forma de volar sobre la mar.
Los paíños son los representantes más pequeños del orden Procellariiformes que incluye también a petreles, albatros y pardelas (Cramp y Simmons, 1977). Los últimos estudios genéticos sitúan al paíño dentro de la gran familia Diomedeidae y, dentro de esta, las distintas especies se agrupan en dos subfamilias, la Hydrobatinae y laOceanitinae. El paíño europeo pertenece al género Hydrobates (Penhallurick y Wink, 2004). Se distinguen dos subespecies: melitensis (Schembri, 1843), presente en la zona mediterránea, y pelagicus, en la costa atlántica europea (Cagnon et al., 2004).
Figura 1. Ejemplar adulto de paíño europeo (© P. López López).
Presenta un tamaño medio de 14-18 cm y una envergadura alar que supera el doble de su longitud corporal. Su peso medio durante la época reproductiva es de unos 28 gramos. A pesar de su aparente fragilidad, soporta sin problemas el oleaje y se le asocia a temporales, por lo que también se le conoce como pájaro de tormenta. Este es el nombre común no sólo en castellano, sino también en catalán (ocell de tempesta), inglés (storm petrel), francés (océanite tempête) e italiano (uccello delle tempeste). Además, en las diferentes comunidades autónomas se le conoce también por otros nombres vernáculos que, en la mayoría de los casos, hacen alusión a su alimentación, anatomía y forma de vida (Tabla 1).
Tabla 1. Algunos de los nombres populares que recibe el paíño.
De cabeza pequeña, sus ojos son oscuros y el pico corto, ganchudo y con grandes narinas (Figura 2). Las patas son negras y cortas con dedos palmeados. Su cola es cuadrada; las alas largas y estrechas. Su cuerpo es de un negro reluciente con una mancha blanca localizada en el obispillo y franjas del mismo color en la cara inferior de las alas (Figura 3). Con los años la tonalidad de las plumas va cambiando del negro a parduzco (Figura 2). Por otra parte, los pollos están cubiertos por plumón grisáceo con algunas calvas (Figura 4).
Figura 2. Detalle de la cabeza del paíño europeo (© M. Gomila).
Figura 3. Paíño europeo (© V. París).
Figura 4. Pollo de paíño europeo (© V. Picorelli).
Tiene un olfato muy desarrollado en consonancia con el tamaño de su bulbo olfatorio. Esta característica es fundamental para identificar otros conspecíficos y localizar su nido (Bonadonna y Sanz-Aguilar, 2012; Dell’Ariccia et al., 2015). Por otra parte, e igual que en otros representantes del orden Procellariiformes, es totalmente reconocible el característico olor intenso de la especie. Como curiosidad destacar que los marineros ingleses consideraban a los paíños portadores de mala suerte, ya que avisaban de la llegada de mal tiempo. Muchos pensaban incluso que eran las almas de marineros fallecidos en el mar.
Vuelo Los paíños vuelan muy próximos a la superficie del mar, alternando elegantes planeos con cortos y rápidos aleteos que recuerdan a los murciélagos (Figura 5). Sus patas suelen colgar rozando la superficie del agua al volar, por lo que parece como si fuesen brincando sobre las olas (Figura 3). Para capturar el alimento vuelan con las patas rozando la superficie marina y picotean buscando las presas que están en la superficie (Albores-Barajas et al., 2011). Pueden volar durante varios días seguidos, recorriendo grandes distancias en busca de alimento (Oppel et al., 2018). Con el inicio del período reproductivo, los paíños realizan vuelos de exhibición, en los que las parejas se suelen perseguir dando vueltas en círculo.
Figura 5. Paíño europeo en vuelo (© V. París).
En tierra, donde sólo se acercan para reproducirse y cuidar de su descendencia, no se mueven con demasiada comodidad, pero imágenes captadas con cámaras infrarrojas de fototrampeo muestran que son capaces de volar dentro de las colonias en total oscuridad.
Biometría y dimorfismo sexual Biometría A continuación se muestran las biometrías de paíños capturados en diferentes colonias españolas (Tablas 2-3).
Tabla 2. Medidas en mm de ala y tarso y en gramos de peso de paíño europeo en dos colonias del Atlántico (Aketx, Bizkaia y Montaña Clara, Canarias) y en varias colonias del Mediterráneo. Se muestran los valores promedio, la desviación estándar y el número de individuos medidos de cada colonia. Se indica con un asterisco aquellas colonias en las que los individuos son capturados con redes. Fuente: datos de Cabrera en Amengual et al. (1999); Montaña Clara, Jacob González-Solís comunicación personal; resto de colonias con datos propios.
Tabla 3. Medidas en mm de cabeza y pico de paíño europeo en dos colonias del Atlántico (Aketx, Bizkaia y Montaña Clara, Canarias) y en varias colonias del Mediterráneo. Se muestran los valores promedio, la desviación estándar y el número de individuos medidos de cada colonia. Se indica con un asterisco aquellas colonias en las que los individuos son capturados con redes. Fuente: datos de Cabrera en Amengual et al. (1999); Montaña Clara, Jacob González-Solís comunicación personal; resto de colonias con datos propios.
Dimorfismo sexual Ambos sexos son idénticos en plumaje (Figura 6) siendo las hembras ligeramente más grandes que los machos (James, 1983; Albores-Barajas et al., 2010). Existen diferentes estudios que proponen una función discriminante para separar ambos sexos. Según James (1983), la longitud del ala y longitud de la cola son las medidas que mejor discriminan los dos sexos, Albores-Barajas et al. (2010) proponen el uso del tamaño de la banda del obispillo y la longitud del ala en un estudio realizado en Marettimo (Mediterráneo) mientras que Jakubas et al. (2014) proponen una función discriminante basada en la longitud del ala y la longitud de la cabeza-pico basado en un estudio que se sexaron 156 individuos en islas Feroe con técnicas moleculares. En la isla de Benidorm no se ha podido obtener una función discriminante fiable en base a biometrías (Tabla 4).
Figura 6. Pareja de paíño europeo en Benidorm (© A. Sanz-Aguilar).
Tabla 4. Medidas en mm de 229 individuos reproductores sexados por técnicas moleculares en la isla de Benidorm. Se muestran los valores promedio, la desviación estándar y el número de individuos medidos (datos propios).
Variación geográfica Se han descrito dos subespecies (Cagnon et al., 2004): H. pelagicus pelagicus (Linnaeus, 1758) que se localiza a lo largo de la costa Europea Atlántica; y la subespecie H. pelagicus melitensis (Schembri, 1843) en el mar Mediterráneo. El análisis filogenético del DNA mitocondrial sugiere que las aves de las poblaciones mediterráneas y atlánticas se separaron al menos hace 350.000 años (Cagnon et al., 2004). Sangster et al. (2012) sugieren que los reproductores mediterráneos deberían reconocerse taxonómicamente, aunque los mantiene a nivel subespecífico a la espera de futuros análisis de vocalizaciones. La diferencia anatómica entre ambas subespecies radica en que las aves del Mediterráneo son ligeramente más grandes y tienen un pico más voluminoso (Hémery y Elbée, 1985; Massa y Catalisano, 1986). También Lalanne sugiere la superior longitud alar y altura de la mandíbula de las aves mediterráneas como la mejor manera de distinguir ambas subespecies (Lalanne et al., 2001). Hydrobates pelagicus melitensis fue descrita por Antonio Schembri (1843) basándose en diferencias de coloración. Estas diferencias también fueron observadas por Ángel Guirao (1859) en aves de Isla Grosa (Murcia) que señala que las aves de los islotes murcianos difieren de las atlánticas en que nunca tienen terminadas de blanco las plumas escapulares ni las remeras secundarias. También Rob et al. (2008) notan que la mayoría de las aves del Mediterráneo muestran un ala más uniformemente negra faltando la banda alar marrón más o menos prominente que muestran las aves del Atlántico. Rob et al (2008) encontraron diferencias en las vocalizaciones comparando grabaciones de 11 aves del Atlántico (Mousa y Skelling) con 12 aves del Mediterráneo (Benidorm y Marettimo) que difieren en la mayor frecuencia alcanzada en una nota (“breath note”) en las aves del Mediterráneo. Anteriormente, Bretagnolle (1998) había comprobado como ambas subespecies mostraban significativamente una mayor respuesta a reclamos de su propio taxón. También existen diferencias fenológicas entre ambos taxones. Las aves del Mediterráneo se reproducen y mudan aproximadamente un mes antes que las aves del Atlántico (Arroyo et al., 2004). Además de las diferencias entre las poblaciones mediterráneas y atlánticas, Jakubas et al. (2014), encuentran un incremento de la longitud alar de oeste a este. A nivel regional también encuentran un incremento de esta variable de sur a norte lo que concuerda con la regla de Bergmann aumentando la longitud alar a medida que disminuye la superficie de mar y la temperatura del aire. Como curiosidad, cuando Ángel Guirao descubrió los paíños de Isla Grosa e Islas Hormigas (Murcia), los consideró como una especie distinta por sus diferencias de coloración con los ejemplares descritos en las islas Feroe y propuso el nombre de Hydrobates stutius por considerarla un ave estúpida que se dejaba coger sin que se defendiera ni huyera.
Muda Los paíños realizan una muda completa del plumaje anualmente (Bolton y Thomas, 2001). La muda comienza en verano, en las colonias de cría, y se prolonga hasta el otoño y comienzos de invierno, ya en sus áreas de invernada (Demongin, 2016). Existen diferencias entre el plumaje juvenil y de adulto, de forma que las plumas de vuelo de los jóvenes suelen estar más apuntadas y son más estrechas que las de los adultos. Los juveniles presentan un plumaje uniforme, donde no se observa una graduación en el desgaste y color de las plumas debido a la progresión de la muda que si muestran los adultos (ver diferencias en las Figuras 7 y 8). Durante la primavera y comienzo del verano, antes de comenzar a mudar, los juveniles de un año de edad (datados como segundo año de calendario; 2 ac en adelante), presentan un plumaje marrón, desgastado que difiere del negro de los adultos. Además, una vez comienza la muda, las nuevas plumas de patrón adulto destacan sobre manera sobre el plumaje juvenil (Figura 7). Al finalizar la muda, en el invierno, un ejemplar de 2ac tendrá un plumaje de adulto que lo hará indistinguible de las aves de mayor edad, de tres años de calendario o más (3ac+). No obstante, algunos ejemplares pueden dejar algunas plumas retenidas, con lo que es posible datar ejemplares de 3ac. En tal caso, en un plumaje adulto, en verano, quedan retenidas una o más plumas juveniles, muy deterioradas. La muda de las plumas de vuelo (primarias- PP-, secundarias-SS- y rectrices-RR-) de los ejemplares de 2ac comienza tres o cuatro semanas antes que la de los adultos, que comienza durante la segunda mitad de la incubación (julio) e incluso más tarde (Demongin, 2016).
Figura 7. Paíño de segundo año de calendario (2ac) capturado el 24 de julio de 2014 en el islote de Aketx, Bizkaia. Se observa que la P1 y la P2 y sus correspondientes coberteras primarias (CCPP) están creciendo y le falta la P3 y la CP3. El plumaje es marrón, muy desgastado, y se observa que las secundarias (SS) son estrechas y apuntadas, sobre todo las internas (© I. Zuberogoitia).
Figura 8. Paíño adulto (3ac+) capturado en nido el 3 de agosto de 2011 con un pollito pequeño, en el islote de Aketx, Bizkaia. Este ejemplar acaba de comenzar la muda, faltándole las PP1 y 2, que aún no asoman. Con mucha atención se puede ver una diferencia en desgaste y color (márgenes y pluma más claros) entre la P3, la primaria más vieja que tiene, y las PP más externas. Esta diferencia es también perceptible en las secundarias (SS), donde se ve que la S1 y la S5 son levemente más viejas que la S4, por ejemplo. Por otro lado, las SS son anchas y con la punta plana (© I. Zuberogoitia).
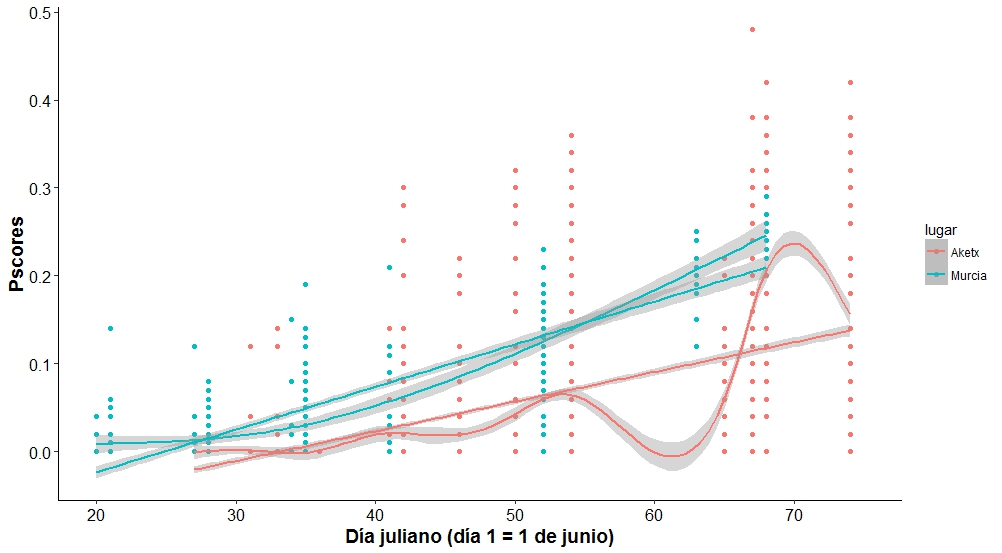
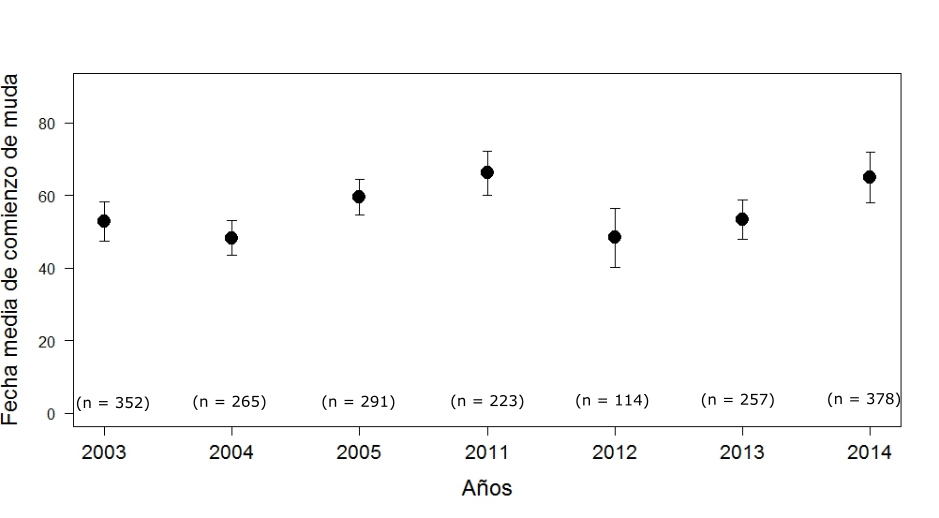
La muda de los paíños comienza con las PP más internas, PP1, y continúa hacia afuera de forma progresiva. Cuando ya han cambiado las cinco o seis primeras PP, entonces comienzan a mudar las secundarias (SS), comenzando en dos centros de muda (SS5 y SS12) y cuando estos ya van avanzando inician un tercer centro de muda en las SS1 (Arroyo et al., 2004). El patrón de muda de las rectrices es más irregular, aunque por lo general comienzan por las RR2, seguidas de las RR1, las RR3 y RR6 y finalmente las RR4 y RR5 suelen ser las últimas en mudarse (Arroyo et al., 2004). La fecha media de inicio de la muda en paíños de la colonia de Benidorm entre 1994 y 1995 fue el 25 de junio para PP, el 21 de julio para SS y tres días más para las RR, mientras que para las colonias de Bizkaia, en 1990 y 1991, fue del 22 de julio para PP (Arroyo et al., 2004). La fecha media de comienzo de muda en Murcia en 2014 fue el 3 de julio (Figura 9; Sallent et al., 2015). La fecha media de inicio de muda de los paíños en Aketx entre 2003 y 2014 fue el 21 de julio (Zuberogoitia et al., 2016). La primera muda activa fue detectada un 2 de julio para Aketx (Zuberogoitia et al., 2016) y un 28 de junio para Mouro (Cantabria) (Gobierno de Cantabria, 2008), mientras que en Murcia (isla Grosa), en el primer muestreo del 20 de junio (2014), el 36% de los paíños (n = 11) ya había comenzado la muda en la P1. Para mediados de agosto (la última visita a la colonia de cría de Aketx) la muda alcanzó la P4, en tres aves hasta la P5 y en tan sólo un ave hasta la P6, y ninguno de los paíños había comenzado a mudar las secundarias. La fecha media de muda fue significativamente diferente entre años, detectándose un retraso significativo (17 días más tarde que en los años 2004 y 2012) en los años 2011 y 2014, asociado a condiciones meteorológicas adversas y la consecución de varios ciclones extratropicales durante el final del invierno y comienzos de primavera (Figura 10; Zuberogoitia et al., 2016). De hecho, en la Figura 9 se observa un descenso notable en la curva de Aketx en el día 65 (4 de agosto), debido a que, en ese año particular, el 32,3% de los ejemplares (n= 96) aún no habían comenzado la muda, y los que ya habían empezado llevaban un retraso de 20 días de media. En septiembre se observan paíños tanto en el Cantábrico como en el Mediterráneo con la muda de PP muy avanzada y con algunas SS cambiadas (básicamente las correspondientes a los centros de muda, ver p.ej. (Gobierno de Cantabria, 2008; Sallent et al., 2015; Juankarandres Blogspot, 2018; Martínez et al., 2019).
Figura 9. Evolución de la muda de los paíños en Aketx según los valores de muda de las primarias (Pscores; de 0 – no muda, a 1 – muda completa) desde el 1 de junio (día 1) hasta el último muestreo de agosto. Los puntos azules muestran los valores de muda en varias colonias de Murcia a lo largo de la temporada de nidificación de 2014; los puntos rojos muestran los valores de muda del islote de Aketx (Bizkaia), acumulados entre 2003 y 2014. Se incluyen las rectas de regresión (± Error estándar) y las curvas de evolución de los valores medios (± Error estándar).
Figura 10. Fecha media e intervalo de confianza (95%) de inicio de la muda de los paíños en Aketx entre 2003 y 2014. La fecha se indica desde el 1 de junio (día 1) hasta el último muestreo de agosto.
Referencias Albores-Barajas, Y.V., Massa, B., Griffiths, K., Soldatini, C. (2010). Sexual dichromatism in Mediterranean Storm Petrel Hydrobates pelagicus. Melitensis. Ardeola, 57: 333-337. Albores-Barajas, Y.V., Riccato, F., Fiorin, R., Massa, B., Torricelli, P., Soldatini, C. (2011) Diet and diving behaviour of European storm petrels Hydrobates pelagicus in the Mediterranean (ssp. melitensis). Bird Study, 58: 208-212. Amengual, J.F., Gargallo, G., Suárez, M., Bonnin, J., González, M., Rebassa, M, McMinn, M. (1999). The Mediterranean Storm Petrel Hydrobates pelagicus melitensis at Cabrera archipiélago (Balearic Island, Spain): breeding moult, biometty and evaluation of the population size by mark and recapture techniques. Ringing & Migration, 19: 181-190. Arroyo, B., Mínguez, E., Palomares, L., Pinilla, J. (2004). The timing and pattern of moult of flight feathers of European Storm-petrel Hydrobates pelagicus in Atlantic and Mediterranean breeding areas. Ardeola, 51: 365-373. Bolton, M., Thomas, R. (2001). Moult and ageing of storm petrels Hydrobates pelagicus. Ringing & Migration, 20: 193-201. Bonadonna, F., Sanz-Aguilar, A. (2012). Kin recognition and inbreeding avoidance in wild birds: the first evidence for individual kin-related odour recognition. Animal Behaviour, 94: 509-513. Bretagnolle, V. (1998). Levels of endemism in Mediterranean Procellariiformes: evidence from morphometrics, behaviour and genetics. In Walmsley, J., Goutner, C, El Hili, A., Sultana, J. (eds) Ecologie des oiseaux marins et gestion intégréedu littoral en méditerranée. IV Symposium Médierranée des oiseaux marins. Les amis des oiseaux y Medmaravis, Tunez. Cagnon, C., Lauga, B., Hémery, G., Mouches, C. (2004). Phylogeographic differentiation of storm petrels (Hydrobates pelagicus) based on cytochrome b mitochondrial DNA variation. Marine Biology, 145: 1257-1264. Cramp, S., Simmons, K. (1977). Birds of the western Palearctic: handbook of the birds of Europe, the Middle East and North Africa. Oxford University Press, New York. Dell’Ariccia, G., Blanc, L., Bonadonna, F., Sanz-Aguilar, A. (2015). Mediterranean storm petrels rely on nest position for homing after migration: a test with artificial nestboxes. Animal Behaviour, 107: 97-104. Demongin, L. (2016). Identification guide to birds in the hand. Cambridge University Press, Cambridge. Gobierno de Cantabria (2008). Seguimiento de la población del paíño europeo Hydrobates pelagicus en Cantabria. Informe inédito de Aves Cantábricas SL para la Consejería de Desarrollo Rural, Ganadería, Pesca y Biodiversidad. Guirao, Á. (1859). Catálogo metódico de las aves observadas en una gran parte de la provincia de Murcia. Memorias de la Real Academia de Ciencias de Madrid. 3ª serie: 2: 540. Hémery, G., Elbée, E. (1985) Descrimination morphologique des populations atlantique et méditerranéene de Pétrel tempête Hydrobates pelagicus. Oiseaux marins nicheurs du Midi et de la Corse. Annales du CROP 2: 63-67 Jakubas, D., Jacubas, W-J., Jensen, J-K. (2014). Body size variation of European Storm Petrels Hydrobates pelagicus in relation to environmental variables. Acta Ornithologica, 49: 71-82. James, P.C. (1983). Storm Petrel Tape Lures: Wich Sex is Attracted? Ringing & Migration, 4: 249-253. Juankarandres Blogspot (2018). http://juankarandres.blogspot.com/search/label/pai%C3%B1o%20europeo. Lalanne Y., Hémery G., Cagnon C., D'Amico F., D'Elbée J., Mouchés C. (2001). Discrimination morphologique des sousespèces d'océanite temp?te: nouveaux résultats pour deux populations mediterranéennes. Alauda, 69: 475–482. Martínez, C., Roscales, J.L., Sanz-Aguilar, A., González-Solís, J. (2019). Inferring the wintering distribution of the mediterranean populations of European storm-petrels (Hydrobates pelagicus melitensis) from stable isotope analysis and observational field data. Ardeola, 66: 13-32. Massa, B., Catalisano, A. (1986). Observations on the Mediterranean Storm Petrel Hydrobates pelagicus at Marettimo Isle. Avocetta 10: 125-127. Oppel, S., Bolton, M., Carneiro, A.P.B., Dias, M.P., Green, J.A., Masello, J.F., Phillips, R.A., Owen, E., Quillfeldt, P., Beard, A., Bertrand, S., Blackburn, J., Boersma, P.D., Borges, A., Broderick, A.C., Catry, P., Cleasby,I., Clingham, E., Creuwels, J., Crofts, S., Cuthbert, R.J., Dallmeijer, H., Davies, D., Davies, R., Dilley, B.J., Dinis, H.A., Dossa, J., Dunn, M.J., Efe, M.A., Fayet, A.L., Figueiredo, L., Pereira Frederico, A., Gjerdrum, C., Godley, B.J., Granadeiro, J.P., Guilford, T., Hamer, K.C., Hazin, C.H., Hedd, A., Henry, L., Hernández-Montero, M., Hinke, J., Kokubun, N., Leat, E., McFarlane Tranquilla, L., Metzger, B., Militão, T., Montrond, G., Mullié, W., Padget, O., Pearmain, E.J., Pollet, I.L., Pütz, K., Quintana, F., Ratcliffe, N., Ronconi, R.A., Ryan, P., Saldanha, S., Shoji, A., Sim, J., Small, C., Soanes, L., Takahashi, A., Trathan, P., Trivelpiece, W., Veen, J., Wakefield, E., Weber, N., Weber, S., Zango, L., Daunt, F., Ito, M., Harris, M.P., Newell, M.A., Wanless, S., González-Solís, J., Croxall, J. (2018). Spatial scales of marine conservation management for breeding seabirds. Marine Policy, 98: 37-46. Penhallurick, J., Wink, M. (2004). Analysis of the taxonomy and nomenclature of the Procellariiformes based on complete nucleotide sequences of the mitochondrial cytochrome b gene. Emu, 104: 125-147. Rob, M., Mullarney, K., Sound Approach (2008). Petrels, night and day. A Sound Approach guide. The Sound Approach, Dorsert. Sallent, A., Ródenas, J.M., Barberá,G.G., Ballestreos, G., García-Castellanos, F.A., Eguía, S., de las Nieves, B. & Aguado, F. (2015). Fenologia de la muda del paíño europeo Hydrobates pelagicus en el sureste Ibérico. V Congreso de la Naturaleza de la Región de Murcia 2015. Libro de resúmenes: 59-60. Murcia del 25 al 28 de noviembre. Sangster, G., Collinson, J. M., Crochet, P-A. Knox, A.G., Parkin, D.T., Votier, S. C. (2012). Taxonomic recomentations for British birds: eight report. Ibis 154: 874-883. Schembri, M. (1843). Catálogo ornitologico del gruppo di Malta. Anglo-Maltese, Malta. Zuberogoitia, I., Zabala, J., Etxezarreta, J., Crespo, A., Burgos, G., Arizaga, J. (2016) Assessing the impact of extreme adverse weather on the biological traits of a European storm petrel colony. Population Ecology, 58: 303-313.
Ana Sanz-Aguilar, Iñigo Zuberogoitia, Ángel Sallent, Fecha de publicación: 4-06-2019 Sanz-Aguilar, A., Zuberogoitia, I., Sallent, Á., Picorelli, V., Navedo, J.G., Garaita, R. (2019). Paíño europeo – Hydrobates pelagicus. En: Enciclopedia Virtual de los Vertebrados Españoles. López, P., Martín, J., González-Solís, J. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||