|
Para más información sobre cada apartado hacer click en:
Portada
Identificación
Estatus de conservación
Distribución
Hábitat
Voz
Movimientos
Ecología trófica
Biología de la reproducción
Interacciones entre especies
Comportamiento
Bibliografía
|
Key words: Storm Petrel, reproduction, reproductive success, demography.
Fenología
La fecha de puesta es un rasgo heredable en esta especie (Sin-Yeon et al., 2012). El periodo de puesta del paíño coincide con picos de disponibilidad de zooplancton, mientras que los pollos nacen cuando la disponibilidad de larvas de peces es máxima (Ramírez et al., 2016). El acoplamiento entre la fenología (fecha de puesta) y la estratificación del agua tiene repercusiones a nivel de éxito reproductor individual y poblacional (Ramírez et al., 2016).
En el área Mediterránea, los paíños comienzan a visitar sus colonias de cría a finales de marzo (Mínguez, 1994). El periodo de puesta se extiende durante unos 3 meses; las primeras puestas se producen a mediados de abril, la mayor parte se concentran entre mediados de mayo y mediados de junio, y las últimas se prolongan hasta la primera quincena de julio (Figura 1; Mínguez, 1994; Sanz-Aguilar et al., 2018a). Se ha observado que las fechas medias y medianas de puesta, así como sus rangos varían en gran medida entre años (Figura 2; Ramírez et al., 2016; Keogan et al., 2018). En las colonias Atlánticas las fechas de puesta son más tardías que en las colonias Mediterráneas, comenzando en mayo (Davis, 1957; Mínguez et al., 1992). Parece ser que la temporada de reproducción de la especie se retrasa conforme las colonias se localizan en zonas más septentrionales (Davis, 1957; Mínguez et al., 1992).
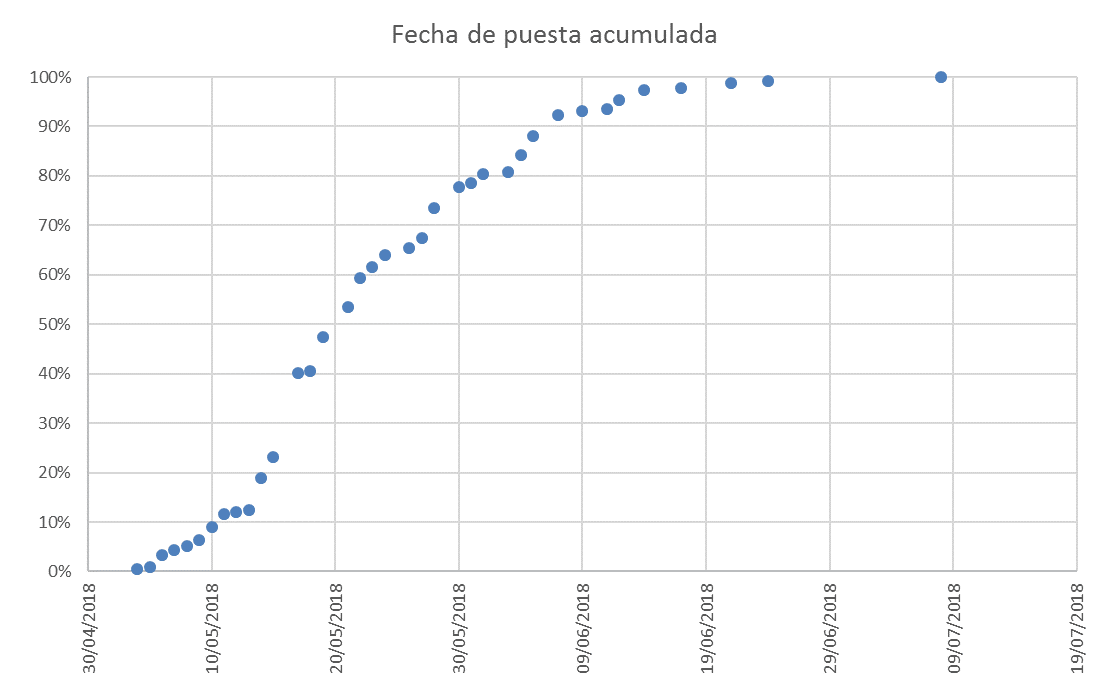
Figura 1. Distribución acumulada de las fechas de puesta del paíño europeo en la Isla de Benidorm en el año 2018 (Sanz-Aguilar et al., 2018a).
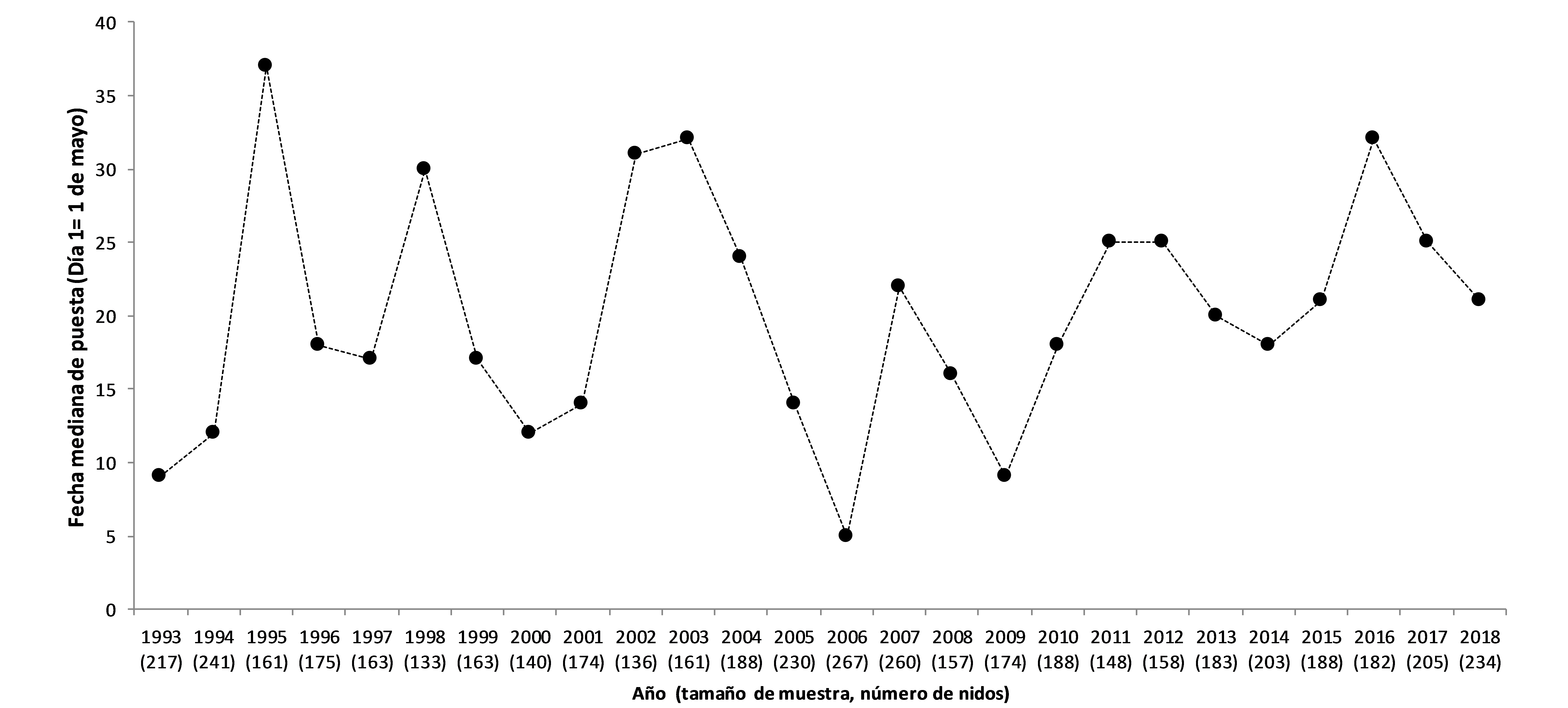
Figura 2. Fechas medianas de puesta del paíño europeo en la isla de Benidorm (Keogan et al., 2018; Sanz-Aguilar et al., 2018a).
Estudios recientes en las colonias de Ogoño, Bizkaia, mediante visitas regulares y cámaras de fototrampeo instaladas de forma continua durante 9 meses, revelaron que los paíños comienzan a acceder a la colonia a finales de febrero (Figura 3) (R. Garaita datos sin publicar). A lo largo del mes de marzo se va incrementando el número de llegadas y salidas a la cueva, y a finales de marzo la colonia se encuentra plenamente ocupada. Las distintas llegadas a la colonia, hasta la puesta del huevo, son para preseleccionar el nido y mantenerlo ocupado. Este periodo es variable, dándose casos de parejas que dedican un mes, hasta parejas que invierten más de 100 días en esta tarea antes de la puesta. Durante el periodo preselectivo, las aves ocupan el lugar del futuro nido de forma intermitente. Según va avanzando el periodo reproductor, se producen las llegadas y salidas para atender las labores propias de la cría. Al final del periodo reproductor, cuando los pollos se van desarrollando y van abandonando la colonia, disminuye el número de movimientos registrados. Además, los progenitores cuyos pollos han volado dejan de retornar a la cueva. Los últimos registros son debidos a las puestas más tardías donde padres y pollos son los últimos en abandonar la colonia.
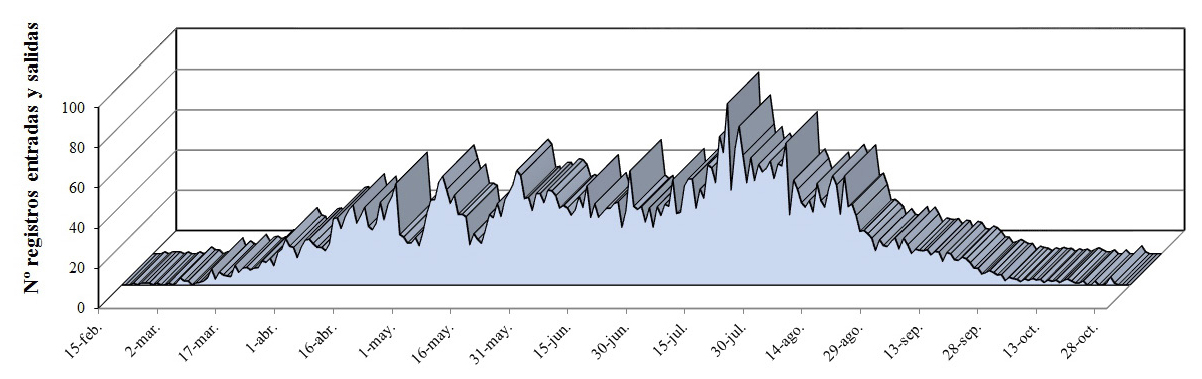
Figura 3. Número de registros de movimientos (entradas y salidas) detectados con cámaras de fototrampeo en la colonia de Ogoño durante el periodo 2013-2016 (a finales de octubre se retiran las cámaras) (R.Garaita datos sin publicar).
Durante el periodo de seguimiento se detectó que los paíños ingieren tierra (geofagia) en los días previos a la puesta (Figura 4) (R. Garaita datos sin publicar). Esta actividad podría estar relacionada con la necesidad de minerales para la formación del huevo. La geofagia acontece en la primera quincena de abril (unos 15 días antes de las primeras puestas) y se detecta hasta la segunda quincena del mes de julio. Fuera de los meses de puestas no se ha detectado este comportamiento.

Figura 4. Paíño ingiriendo tierra en la colonia de Ogoño (© R. Garaita).
Las primeras puestas se producen en la segunda quincena de abril (p. ej., 21 abril 2016) y las sucesivas se alargan a lo largo del verano, detectándose puestas tardías a mediados de agosto (Figura 5). Aunque los primeros pollos nacen a finales de mayo-primeros de junio, la mayoría lo hace a lo largo del mes de julio. Los pollos de las escasas puestas tardías de agosto que consiguen eclosionar nacen en septiembre (Figura 5). Los primeros pollos vuelan a mediados de agosto, la mayoría a lo largo del mes septiembre y los últimos, de las puestas tardías, volarían a finales de noviembre-primeros de diciembre (Figura 6; Garaita, 2016).
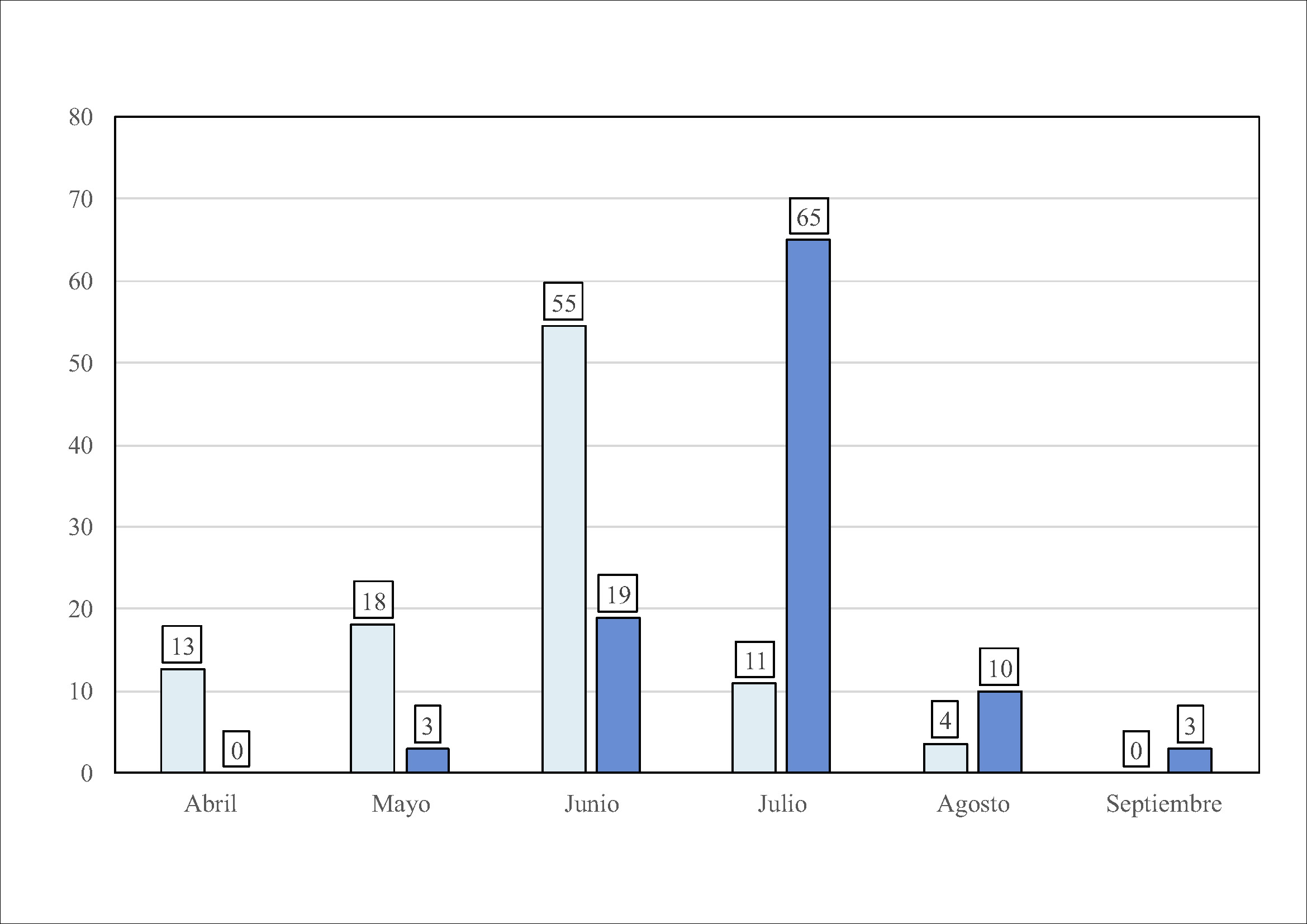
Figura 5. Porcentaje mensual de puestas (azul claro) y eclosiones (azul oscuro) en la colonia de Ogoño durante el periodo 2013-2016 (R.Garaita datos sin publicar).

Figura 6. Pollo de paíño procedente de una puesta tardía estimada entre el 15-18 de agosto de 2016. Se estima que nació entre el 25-28 de septiembre. En el momento que se tomó la foto (28 de octubre) tendría unos 30 días y presentaba buena condición corporal aparente, por lo que de sobrevivir abandonaría el nido a finales de noviembre o primeros de diciembre (© R. Garaita).
Reclutamiento
Generalmente, el reclutamiento en la fracción reproductora de la población se produce entre los 3 y 5 años de edad (entre 4ac y 6ac; Okill y Bolton, 2005; Sanz-Aguilar et al., 2009a). Existen importantes diferencias en la edad de la primera reproducción (reclutamiento) entre diferentes colonias de la especie (Tabla 1, Sanz-Aguilar et al., 2009a). En Benidorm nunca se ha observado un reclutamiento de paíños menores a 3 años de edad (4ac). Sin embargo, en s’Espartar (Ibiza) se constató la reproducción de un paíño de 2 años de edad (3ac, Sanz-Aguilar et al., 2018b). En la colonia de Marettimo (Sicilia, Italia) se observó que unos pocos individuos son capaces de reclutar en la fracción reproductora de la población su primer año de vida (2ac, LoValvo y Massa, 2000; Sanz-Aguilar et al., 2009a). La aparición de jóvenes (2ac) en la isla de Aketx (Bizkaia) está aparentemente relacionada con eventos que afectan la dinámica de reproducción de las aves adultas, tales como la marea negra del Prestige o fenómenos meteorológicos extremos (Tabla 2; Zabala et al., 2011; Zuberogoitia et al., 2016).
Tabla 1. Proporción de reproductores del paíño europeo para cada clase de edad en Marettimo (Sicilia, Italia) y las dos colonias de la Isla de Benidorm (Alicante, España) (Sanz-Aguilar et al., 2009a). Para la edad se muestran los años de vida y, entre paréntesis, los años de calendario (ac).
Edad |
Marettimo |
Benidorm (C1) |
Benidorm (C2) |
1 (2ac) |
7% |
0% |
0% |
2 (3ac) |
15% |
0% |
0% |
3 (4ac) |
65% |
11% |
97% |
4 (5ac) |
65% |
92% |
100% |
5 (6ac) |
100% |
100% |
100% |
Tabla 2. Proporción de juveniles (2ac) capturados durante los muestreos de julio y agosto en el islote de Aketx (Zuberogoitia et al., 2014).
Año |
Proporción de Juveniles (%) |
2003 |
2,97 |
2004 |
0,35 |
2005 |
0,35 |
2006 |
2,3 |
2011 |
4,46 |
2012 |
0 |
2013 |
0,78 |
2014 |
5,82 |
Éxito reproductor
El paíño europeo pone un único huevo de grandes dimensiones (6,77±0,6 gramos; 23.5%) con respecto al peso medio corporal de un adulto (A.Sanz-Aguilar datos sin publicar). En ocasiones excepcionales, si la puesta fracasa al principio de la temporada de cría, son capaces hacer una puesta de reposición (Mínguez, 1997). El huevo es incubado por ambos sexos durante unos 40 días, en turnos de incubación medios de 2 días, pero que pueden variar entre 1 y 5 días (Davis, 1957; Scott, 1970; Mínguez, 1998).
Tras la eclosión del huevo, el pollo no es capaz de termorregular por sí sólo, por lo que los adultos lo “empollan”, es decir, permanecen durante todo el día sobre él, durante la primera semana de vida (Figura 7; Mínguez y Oro, 2003). A partir de entonces, tan sólo es visitado por la noche por ambos progenitores, de forma intermitente, para proporcionarle alimento. En torno a los 54 días de edad, los pollos alcanzan el 150% del peso de los adultos (Mínguez, 1996). La frecuencia de las cebas va disminuyendo y en torno a los 63-70 días de vida los pollos abandonan las colonias (Davis, 1957; Bolton, 1995a, b; Mínguez, 1996).

Figura 7. Paíño adulto con su pollo en la colonia de s’Espartar, Ibiza (© M. Viñas).
En algunas colonias atlánticas se ha descrito intermitencia facultativa en la reproducción, es decir, la existencia de años sabáticos entre temporadas reproductoras (Hemery et al., 1986). En el islote de Aketx (Bizkaia) el porcentaje de nidos ocupados difiere entre años en función de factores meteorológicos y la existencia de condiciones adversas (Tabla 3; Azkona et al., 2006; Zabala et al., 2011; Zuberogoitia et al., 2016). Sin embargo, en otras colonias, como en la isla de Benidorm, la mayoría de las parejas se reproducen anualmente (Sanz-Aguilar et al., 2008). El éxito reproductor de la especie muestra variaciones interanuales y espaciales (Tabla 3; Tavecchia et al., 2008). A su vez, en colonias donde los nidos se encuentran relativamente expuestos (p.ej. nidos bajo la vegetación), las condiciones ambientales (como los periodos de abundantes lluvias) pueden reducir el éxito reproductor (Cadiou et al., 2010). Las interacciones con otras especies de aves pueden asimismo afectar a su éxito reproductor, tanto por depredación directa de adultos o de pollos como por destrucción de su hábitat de cría (Cadiou et al., 2010; Libois et al., 2012; Sanz-Aguilar et al., 2009b).
Tabla 3. Éxito reproductor medio del Paíño europeo (se incluye entre paréntesis el número de nidos contolados) en la Isla de Benidorm (colonias 1 y 2, Alicante), Isla Mitjana (Alicante), Isla Tabarca (Alicante), Islas Columbretes (Castellón), Isla s’Espartar (Ibiza) y en el islote de Aketx (Bizkaia) y Ogoño (Bizkaia) donde los nidos monitorizados son una pequeña fracción, accesible, de la colonia. Referencias: García y Martínez, 2008; Garaita y Del Villar, 2008; García, 2009; Generalitat Valenciana, 2013; Garaita, 2016; Zuberogoitia et al., 2016; Sanz-Aguilar et al., 2018a, 2018b.
Año |
Benidorm C1 |
Benidorm C2 |
Mitjana |
Tabarca |
Columbretes |
Espartar |
Aketx |
Ogoño |
1993 |
0,77 (209) |
0,45 (64) |
|
|
|
|
|
|
1994 |
0,59 (206) |
0,46 (59) |
|
|
|
|
|
|
1995 |
0,51 (174) |
0,47 (47) |
|
|
0,78 (9) |
|
|
|
1996 |
0,54 (140) |
0,24 (41) |
|
|
0,13 (16) |
|
|
|
1997 |
0,48 (142) |
0,63 (35) |
|
|
0,40 (15) |
|
|
|
1998 |
0,35 (142) |
0,53 (38) |
|
|
0,40 (15) |
|
|
|
1999 |
0,48 (145) |
0,59 (56) |
|
|
|
|
|
|
2000 |
0,44 (144) |
0,45 (64) |
|
|
|
|
|
|
2001 |
0,50 (113) |
0,59 (70) |
|
|
|
|
|
|
2002 |
0,44 (84) |
0,49 (49) |
|
|
0,37 (8) |
|
|
|
2003 |
0,55 (104) |
0,53 (64) |
|
|
|
|
0,17 (12) |
|
2004 |
0,60 (114) |
0,56 (77) |
|
|
|
|
1,00 (6) |
|
2005 |
0,62 (141) |
0,61 (97) |
|
|
|
|
0,93 (15) |
|
2006 |
0,67 (175) |
0,55 (109) |
|
|
0,5 (2) |
|
|
0,50 (26) |
2007 |
0,70 (174) |
0,73 (94) |
|
|
|
|
|
0,44 (34) |
2008 |
0,66 (102) |
0,72 (67) |
|
|
|
0,58 (¿?) |
|
0,63 (32) |
2009 |
0,50 (110) |
0,54 (67) |
0,60 (20) |
0,50 (14) |
|
0,64 (28) |
|
|
2010 |
0,72 (118) |
0,75 (73) |
|
0,11 (9) |
|
|
|
|
2011 |
0,65 (108) |
0,68 (68) |
0,73 (49) |
0,88 (16) |
|
|
|
|
2012 |
0,55 (103) |
0,69 (68) |
|
0,36 (5) |
|
|
0,92 (12) |
0,61 (49) |
2013 |
0,40 (123) |
0,48 (94) |
|
0,72 (11) |
|
|
0,92 (12) |
0,71(38) |
2014 |
0,55 (103) |
0,67 (104) |
|
|
|
0,45 (184) |
0,13 (8) |
0,52 (42) |
2015 |
0,49 (101) |
0,65 (110) |
|
|
|
0,49 (163) |
|
0,71 (31) |
2016 |
0,39 (89) |
0,67 (105) |
|
|
|
0,44 (192) |
|
0,78 (40) |
2017 |
0,54 (111) |
0,61 (100) |
|
|
|
0,45 (163) |
|
|
2018 |
0,61 (129) |
0,75 (110) |
|
|
|
0,44 (171) |
|
|
2018 |
0,61 (129) |
0,75 (110) |
|
|
|
0,44 (171) |
|
|
En nidos naturales los huevos pueden ser dañados por pequeñas piedras que se desprendan o se encuentren en la superficie del nido (Bolton et al., 2004). Este hecho, unido a la reducción en las interferencias como las peleas territoriales entre individuos en las colonias y la mayor protección ante depredadores, hace que el éxito de eclosión y reproductor de los individuos que crían en nidales artificiales sea normalmente superior (Libois et al., 2012).
Gracias al uso de cámaras de fototrampeo, en la colonia de Ogoño, durante el periodo 2013-2016, se determinaron algunas causas del fracaso reproductor, tales como el desplazamiento y abandono de los huevos fuera de nido, incubaciones interrumpidas en distintos estadios, la muerte de pollos divagantes (que se aventuran fuera del nido) y/o abandonados (Garaita, 2012, 2013, 2014, 2015, 2016).
La supervivencia de los pollos en nido es generalmente elevada, siendo el promedio de los últimos 26 años en la isla de Benidorm de 0,85 (Mínguez y Oro, 2003; Sanz-Aguilar et al., 2018a). El periodo crítico de supervivencia de los pollos en el nido son los primeros días de vida, es decir, el periodo en que los progenitores “empollan” a su descendiente (Mínguez y Oro, 2003). Las diferencias interanuales observadas en la supervivencia de los pollos sugieren que la cantidad de cuidado parental, en términos de alimentación y aporte de calor, podría estar condicionada por la disponibilidad de alimento durante el periodo de “empollamiento” (Mínguez y Oro, 2003). Datos recientes de la colonia de s’Espartar en Ibiza apuntan a que las interacciones con parásitos (garrapatas) pueden afectar también a la supervivencia de los pollos durante sus primeras semanas de vida (Sanz-Aguilar et al., 2018b), como se había sugerido en trabajos anteriores (Merino et al., 1999).
A nivel individual, la experiencia reproductora, calidad y edad del individuo afectan a sus parámetros reproductores (Sanz-Aguilar et al., 2008; Sanz-Aguilar 2009a; Hernández et al., 2017). El éxito reproductor aumenta conforme aumenta la edad y, sobre todo, con la experiencia reproductora del individuo (Sanz-Aguilar et al., 2008; Sanz-Aguilar 2009a; Hernández et al., 2017). Sin embargo, a edades muy avanzadas se observa un descenso en el éxito reproductor, probablemente asociado con la senescencia (Hernández et al., 2017). Existe una mayor probabilidad de tener éxito en la reproducción del año siguiente para individuos exitosos que para individuos que fracasan, lo que indica un efecto de la calidad del individuo en el éxito reproductor (Sanz-Aguilar et al., 2008).
Supervivencia
El Paíño europeo es una especie longeva. Por el momento el récord de longevidad se encuentra en >33 años (European Longevity Records, Euring). El paíño más longevo capturado en las colonias cantábricas fue anillado de adulto (3ac+) en 1991 y recapturado criando en 2013, con 25 ac+ (Aketx, Bizkaia, Oficina de Anillamiento de Aranzadi). En Benidorm, hasta la fecha, la edad máxima que han alcanzado los paíños marcados como pollos que han vuelto a criar a la isla son los 21 años (22 ac). Cabe destacar que en 2017 se recapturó un reproductor que fue anillado también como reproductor (3ac+) en el año 1995 que, hasta la fecha, sería el récord de edad en las colonias ibéricas mediterráneas 25 ac+; 23 años criando).
Una de las características comunes en todas las colonias de paíño estudiadas es la presencia de individuos “transeúntes” (Sanz-Aguilar et al., 2010). En el caso de reproductores, se trata de individuos que se reproducen en tan sólo una sola ocasión y desaparecen, lo que podría ser el resultado de un coste de la primera reproducción (Sanz-Aguilar et al., 2008). En la isla de Benidorm, se ha observado que este coste de la primera reproducción (menor supervivencia) se da principalmente en hembras (Sanz-Aguilar et al., 2012), y en individuos de baja calidad que presentan menor éxito en la reproducción (Sanz-Aguilar et al., 2008). En estudios realizados mediante captura en redes se observa una mayor proporción de transeúntes capturados, debido a la captura de individuos jóvenes prospectores (Sanz-Aguilar et al., 2010). Estudios recientes indican que los individuos prospectores presentan un peso ligeramente inferior al de individuos reproductores (Cadiou, 2016). La vascularización de la placa incubatriz no representa una señal clara de estatus reproductor, ni tampoco la muda o las medidas biométricas (Cadiou, 2016). Por lo tanto, resulta complicado poder asignar el estatus reproductor de individuos capturados mediante redes (Sanz-Aguilar et al., 2010; Cadiou, 2016). Estimas obtenidas en el archipiélago de Molene (Bretaña) indican que alrededor del 85% de las aves capturadas en redes (y un 75% de los nuevos anillamientos) serían muy probablemente individuos prospectores (Cadiou, 2016). Sin embargo, en el Mediterráneo, donde la población de paíños es más pequeña que en el Atlántico, la proporción de prospectores podría ser menor (Tabla 4).
La supervivencia adulta de los individuos residentes es variable tanto a nivel espacial (Tabla 4) como temporal (Tavecchia et al., 2008). Puede verse afectada por presencia de depredadores, por factores ambientales en las zonas de invernada, durante la migración o en las etapas previas a la reproducción (Oro et al., 2005; Sanz-Aguilar et al., 2009b; Soldatini et al., 2014; Zuberogoitia et al., 2016; Matovic et al., 2017). También se ha observado una mayor supervivencia de individuos que crían en cajas nido con respecto a individuos que se reproducen en nidos naturales, probablemente debida a una mayor protección frente a depredadores (Libois et al., 2012).
Tabla 4. Supervivencia media e intervalos de confianza 95% del Paíño europeo en diferentes colonias españolas (datos propios). En el caso de la Isla de Benidorm (colonias 1 y 2, Alicante) y s’Espartar (Ibiza) los datos corresponden a individuos reproductores. En el resto de colonias los datos corresponden a capturas en red de individuos reproductores y no-reproductores. Se detalla a su vez la proporción de individuos transeúntes entre los individuos capturados/anillados por primera vez en cada zona de estudio. Fuentes: a Gobierno de Cantabria, 2018; b Zuberogoitia et al., 2016.
Período |
Colonia |
1º reproducción obs. |
Residentes |
% Transeúntes |
1993-2018 |
C1 Benidorm, Alicante |
0,72 (0,67-0,77) |
0,83 (0,81-0,85) |
13% |
1994-2018 |
C2 Benidorm, Alicante |
0,74 (0,67-0,80) |
0,88 (0,85-0,90) |
16% |
2014-2018 |
S’Espartar, Ibiza |
0,72 (0,64-0,78) |
0,87 (0,77-0,93) |
17% |
2004-2017 |
Palomas, Murcia |
- |
0,83 (0,77-0,88) |
51% |
2004-2014 |
Hormigas, Murcia |
- |
0,90 (0,86-0,93) |
36% |
2007-2015 |
Grosa, Murcia |
- |
0,87 (0,76-0,94) |
65% |
2006-2018a |
Mouro, Cantabria |
- |
0,86 (0,83-0,88) |
63% |
1993-2014b |
Aketx, Bizkaia |
- |
0,81 (0,77-0,85) |
75% |
A nivel individual, factores como la edad, experiencia reproductora y calidad del individuo influyen en la supervivencia. La probabilidad de supervivencia aumenta con la edad y la experiencia reproductora del individuo, y resulta superior en el caso de individuos de alta calidad (que se reproducen exitosamente) (Sanz-Aguilar et al., 2008; Hernández et al., 2017). Podría existir un ligero descenso de la supervivencia entre las clases de edad más avanzadas debido a procesos de senescencia (Hernández et al., 2017).
Dinámica poblacional
Los censos de nidos observables en la isla de Benidorm (Alicante) y s’Espartar (Ibiza) muestran tendencias anuales diferenciales a nivel de colonia (Figuras 8-9; Sanz-Aguilar et al., 2018a, 2018b). Aunque se observan variaciones anuales, no se puede indicar una tendencia generalizada al declive o crecimiento en las diferentes poblaciones estudiadas. Por ejemplo, durante los últimos 26 años, en la colonia 1 de Benidorm se observa una tasa neta de declive anual promedio de -0,32%, mientras que la colonia 2 ha experimentado una tasa neta de crecimiento anual promedio de +3,16% (Figuras 8-9). Ante valores de supervivencia adulta inferiores a 0,8 (80%), las poblaciones podrían mostrar tasas de crecimiento poblacional negativo, tal y como se observaba en la colonia 1 de la isla de Benidorm antes de que se realizara un descaste selectivo de gaviotas especializadas en depredar paíños en el año 2004 (Sanz-Aguilar et al., 2009a, 2009b). De hecho, la colonia mostró tasas de crecimiento poblacional positivo tras la eliminación de gaviotas especialistas (Tenan et al., 2014).
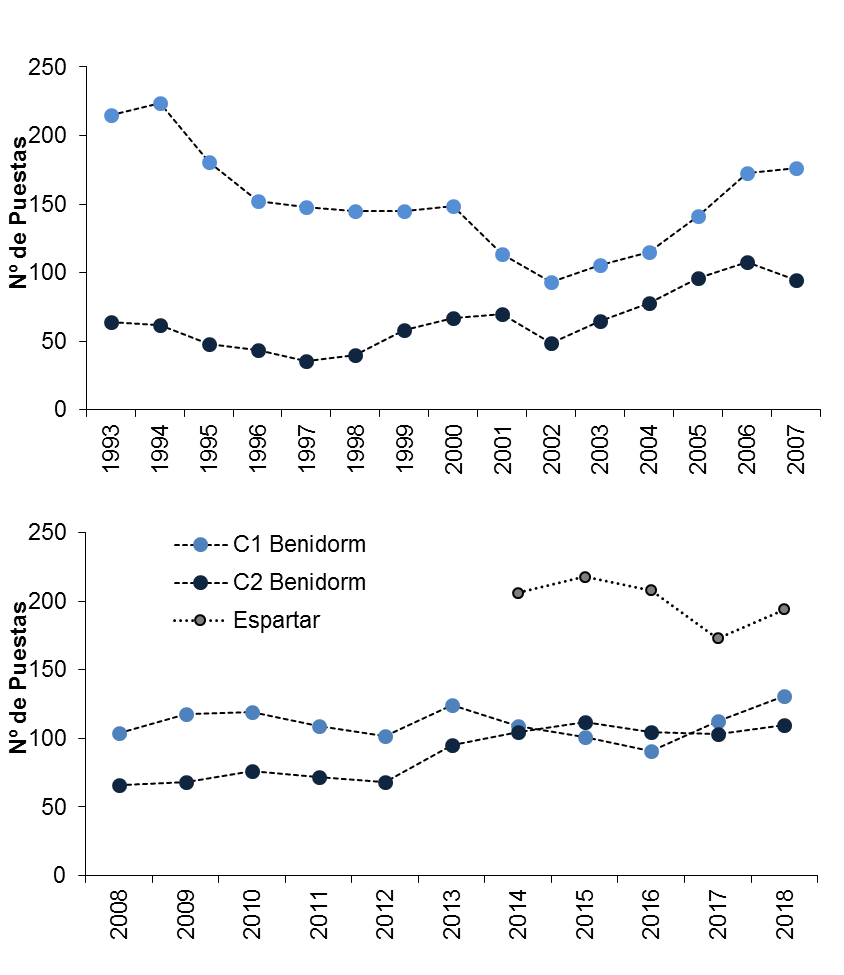
Figura 8. Evolución anual del número de puestas (parejas reproductoras) de paíño europeo en nidos controlados de la Isla de Benidorm (colonias 1 y 2, Alicante) y s’Espartar (Ibiza). En el caso de la Isla de Benidorm el protocolo de seguimiento se redujo el año 2008 y por lo tanto el número de nidos en seguimiento disminuye dicho año. Los datos son comparables para los años presentes en cada uno de los paneles de la Figura (Sanz-Aguilar et al., 2018a, 2018b).
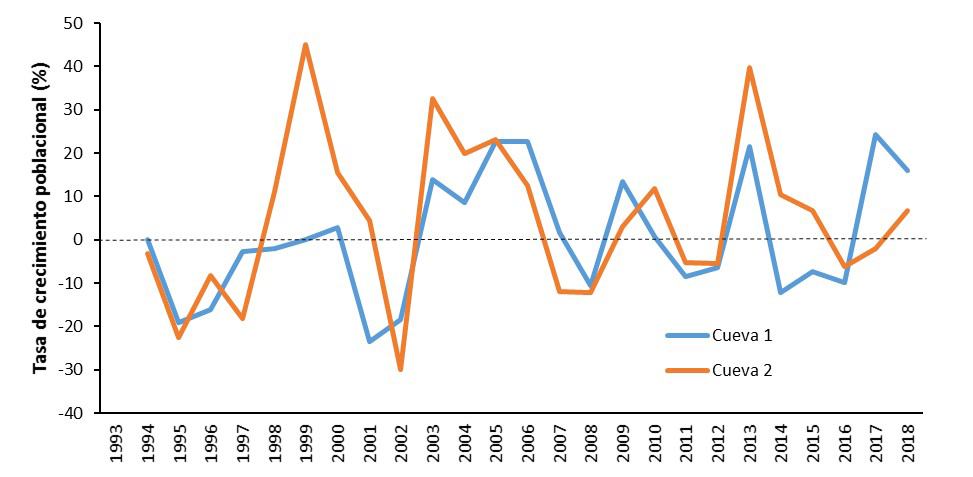
Figura 9. Tasas de crecimiento poblacional del paíño en base al número de nidos activos (con puesta) detectados anualmente en las dos colonias estudiadas en la Isla de Benidorm (1993-2018). La línea de puntos indica la estabilidad, crecimiento/decrecimiento=0.
En cuanto a las poblaciones cantábricas, no hay datos concretos de tamaño de población debido al difícil acceso de las colonias y lo repartido que están los nidos en grietas, fisuras y agujeros dispersos por paredes y acantilados. Los cálculos de tendencias se extraen de las capturas, debiendo tomar las correspondientes reservas, dado que en ellas se incluyen reproductores y transeúntes. Atendiendo a estos datos, no se observa una clara tendencia en el número de capturas en el islote de Aketx, Bizkaia (Figura 10; Zuberogoitia et al., 2014).
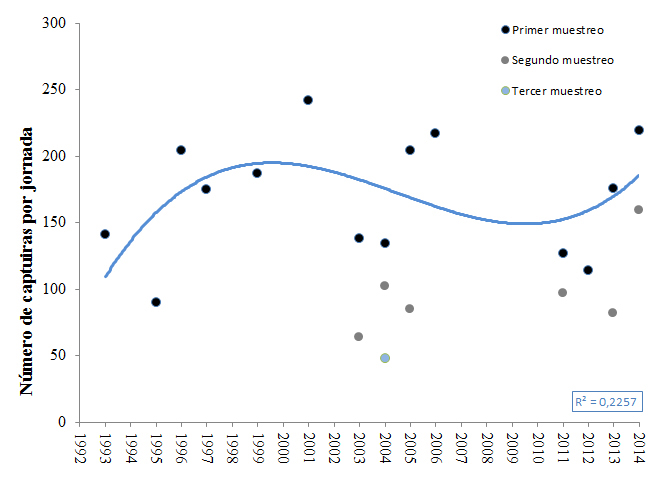
Figura 10. Evolución de las capturas a lo largo de las últimas dos décadas en el islote de Aketx, Bizkaia. El primer muestreo se realizaba durante la última semana de junio y primeros días de julio, el segundo a finales de julio y primera quincena de agosto y el tercer muestreo a mediados de agosto. Se muestra una línea logarítmica ajustada a los puntos del primer muestreo. Los muestreos fueron siempre iguales (mismo número de redes y misma posición) y sin usar reclamos.
Referencias
Azkona, A., Zuberogoitia, I., Martinez, J.A., Etxezarreta, J., Iraeta, A., Castillo, I., Zabala, J., Hidalgo, S. (2006). Short-term effects of the prestige oil spill on a colony of European storm-petrels Hydrobates pelagicus. Acta Zoologica Sinica, 52: 1042-1048.
Bolton, M. (1995a). Experimental evidence for regulation of food delivey to storm petrel, Hydrobates pelagicus, nestlings: the role of chick body condition. Animal Behaviour, 50: 231-236.
Bolton, M. (1995b). Food delivery to nestling storm petrels: limitation or regulation? Functional Ecology, 9: 161-170.
Bolton, M., Medeiros, R., Hothersall, B., Campos, A. (2004). The use of artificial breeding chambers as a conservation measure for cavity-nesting procellariiform seabirds: a case study of the Madeiran storm petrel (Oceanodroma castro). Biological Conservation, 116: 73-80.
Cadiou, B. (2016). New data about the secret life of the wandering prebreeding European storm petrels at colonies. Poster. Seabird Group 13th International Conference, 6-9 September 2016, Edimburgh, UK.
Cadiou, B., Bioret, F., Chenesseau, D. (2010). Response of breeding European Storm Petrels Hydrobates pelagicus to habitat change. Journal of Ornithology, 151: 317-327.
Davis, P. (1957). The breeding of the Storm Petrel. British Birds, 50: 85-101.
Garaita, R. (2012). El Paíño europeo (Hydrobates pelagicus) en Urdaibai. Informe 2012. Departamento de Medio Ambiente de la Diputación Foral de Bizkaia. Informe inédito.
Garaita, R. (2013). El Paíño europeo (Hydrobates pelagicus) en Urdaibai. Informe 2013. Departamento de Medio Ambiente de la Diputación Foral de Bizkaia. Informe inédito.
Garaita, R. (2014). El Paíño europeo (Hydrobates pelagicus) en Urdaibai. Informe 2014. Departamento de Medio Ambiente de la Diputación Foral de Bizkaia. Informe inédito.
Garaita, R. (2015). El Paíño europeo (Hydrobates pelagicus) en Urdaibai. Informe 2015. Biodibertsitatea eta Paisaia Zaintzeko Atala / Sección de Biodiversidad y Paisaje. Iraunkortasuna eta Ingurune Naturala Zaintzeko Saila / Departamento de Sostenibilidad y Medio Natural. Bizkaiko Foru Aldundia / Diputación Foral de Bizkaia. Informe inédito.
Garaita, R. (2016). El paíño Europeo Hydrobates pelagicus en Urdaibai. Informe 2016. Biodibertsitatea eta Paisaia Zaintzeko Atala / Sección de Biodiversidad y Paisaje. Iraunkortasuna eta Ingurune Naturala Zaintzeko Saila / Departamento de Sostenibilidad y Medio Natural. Bizkaiko Foru Aldundia / Diputación Foral de Bizkaia. Informe inédito.
Garaita, R., Del Villar, J. (2008). El paíño europeo (Hydrobates pelagicus) en Urdaibai. Gobierno Vasco. Departamento de Medio Ambiente y Ordenación del Territorio. Informe inédito.
García, D. (2009). Éxito reproductor y campaña de anillamiento del paíño europeo (Hydrobates pelagicus) en el islote de s’Espartar 2009. Reserves naturals d’es Vedrà, es Vedranell i els illots de Ponent. Conselleria de Medi Ambient. Govern de les Illes Balears. Informe inédito.
García, D., Martínez, O. (2008). Censo de los efectivos reproductores del paíño europeo (Hydrobates pelagicus) y uso del hábitat en los islotes de les Reserves Naturals d’es Vedrà, es Vedranell i els illots de Ponent. Conselleria de Medi Ambient. Govern Balear. Informe inédito.
Generalitat Valenciana (2013). Informe sobre el seguimiento del Paíño europeo Hydrobates pelagicus en la Comunitat Valenciana. Año 2013. Generalitat Valenciana. Informe inédito.
Gobierno de Cantabria (2018). Memoria Técnica Justificativa del Plan de Recuperación del paíño europeo Hydrobates pelagicus en Cantabria. Asistencia Técnica de Aves Cantábricas SL a la Dirección General del Medio natural de la Consejería de Medio Rural, Pesca y Alimentación del Gobierno de Cantabria.
Hemery, G., Delbee, E., Terrasse, J.F. (1986). Regulation of a storm-petrel Hydrobates pelagicus population by intermittent breeding. Comptes Rendus de l’Academie des Sciences Serie Iii-Sciences de la Vie-Life Sciences, 303: 353-356.
Hernández, N., Oro, D., Sanz-Aguilar, A. (2017). Environmental conditions, age, and senescence differentially influence survival and reproduction in the Storm Petrel. Journal of Ornithology, 158: 113-123.
Keogan, K., Daunt, F., Wanless, S., Phillips, R.A., Walling, C.A., Agnew, P., Ainley, D.G., Anker-Nilssen, T., Ballard, G., Barrett ,R.T., Barton, K.J., Bech, C., Becker, P., Berglund, P-A ., Birkhead T., Bollache, L., Bond, A., Bouwhuis, S., Bradley, R., Burr, Z .,Camphuysen, K., Catry, P., Chiaradia, A., Christensen-Dalsgaard, S., Cuthbert, R .,Dehnhard, N., Descamps, S., Diamond, T., Divoky, G., Drummond, H., Dugger, KM., Dunn, MJ .,Emmerson, L., Erikstad, KE., Fort, J., Fraser, W., Gaston, A., Genovart, M., Gilg, O., González-Solís, J., Gremillet, D., Hansen, J., Hanssen, S. A., Harris, M., Hedd, A., Hinke, J., Igual, J. M., Jahncke, J ., Jones, I., Kappes, P.J., Lang, J., Langset, M., Lescroël, A., Lorentsen, S.H., Lyver, P.O’B., Mallory, M., Moe, B., Montevecchi, W.A., Monticelli, D., Mostello, C., Newell, M., Nicholson, L ., Nisbet, I., Olsson, O., Oro, D., Pattison, V., Poisbleau, M., Pyk, T., Quintana, F., Ramos, J., Ramos, R., Reiertsen, TK., Rodríguez, C., Ryan, P., Sanz-Aguilar, A., Schmidt, N.M., Shannon, P., Sittler, B., Southwell, C., Surman, C., Svagelj, W.S., Trivelpiece, W., Warzybok, P., Watanuki, Y., Weimerskirch, H., Wilson, P. R., Wood, A. G., Phillimore. A. B., Lewis. S. (2018) Global phenological insensitivity to shifting ocean temperatures among seabirds. Nature Climate Change, 8:313-318.
Libois, E., Gimenez, O., Oro, D., Mínguez, E., Pradel, R., Sanz-Aguilar, A. (2012). Nest boxes: A successful management tool for the conservation of an endangered seabird. Biological Conservation, 155: 39-43.
LoValvo, F., Massa, B. (2000). Some aspects of the population structure of storm petrels Hydrobates pelagicus breeding on a Mediterranean island. Ringing & Migration, 20: 125-128.
Matovic, N., Cadiou, B., Oro, D., Sanz-Aguilar, A. (2017). Disentangling the effects of predation and oceanographic fluctuations in the mortality of two allopatric seabird populations. Population Ecology, 59: 225-238.
Merino, S., Mínguez, E., Belliure, B. (1999). Ectoparasite effects on nestling European storm-petrels. Waterbirds, 22: 297-301.
Mínguez, E. (1994). Censo, cronología de puesta y éxito reproductor del paíño común (Hydrobates pelagicus) en la Isla de Benidorm (Alicante E de España). Ardeola, 41 :3-11.
Mínguez, E. (1996). Nestling feeding strategy of the British storm-petrel Hydrobates pelagicus in a Mediterranean colony. Journal of Zoology, 239: 633-643.
Mínguez, E. (1997) Evidence of occasional re-laying in the British Storm-petrel (Hydrobates pelagicus). Colonial Waterbirds, 20: 102-104.
Mínguez, E. (1998). The costs of incubation in the British Storm-petrel: an experimental study in a single-egg layer. Journal of Avian Biology, 29: 183-189.
Mínguez, E., Elizondo, R., Balerdi, M., Saban, P. (1992). Statut, distribution, taille de la population et phénologie de la reproduction du Pétrel tempête Hydrobates pelagicus dans la communauté autonome basque (Espagne). L’Oiseau et RFO, 62: 234-246.
Mínguez, E., Oro, D. (2003.) Variations in nest mortality in the European storm petrel Hydrobates pelagicus. Ardea, 91: 113-117.
Okill, J.D., Bolton, M. (2005). Ages of Storm Petrels Hydrobates pelagicus prospecting potential breeding colonies. Ringing & Migration, 22: 205-208.
Oro, D., De León, A., Mínguez, E., Furness, R.W. (2005). Estimating predation on breeding European storm-petrels (Hydrobates pelagicus) by yellow-legged gulls (Larus michahellis). Journal of Zoology, 265: 421-429.
Ramírez, F., Afán, I., Tavecchia, G., Catalán, I.A., Oro, D., Sanz-Aguilar, A. (2016). Oceanographic drivers and mistiming processes shape breeding success in a seabird. Proceedings of the Royal Society Series B, 283: 20152287
Sanz-Aguilar, A., Tavecchia, G., Pradel, R., Mínguez, E., Oro, D. (2008). The cost of reproduction and experience-dependent vital rates in a small petrel. Ecology, 89: 3195-3203.
Sanz-Aguilar, A., Massa, B., LoValvo, F., Oro, D., Mínguez, E., Tavecchia, G. (2009a). Contrasting age-specific recruitment and survival at different spatial scales: a case study with the European storm petrel. Ecography, 32: 637-646.
Sanz-Aguilar, A., Martínez-Abraín, A., Tavecchia, G., Mínguez, E., Oro, D. (2009b). Evidence-based culling of a facultative predator: efficacy and efficiency components. Biological Conservation, 142: 424-431.
Sanz-Aguilar, A., Tavecchia, G., Mínguez, E., Massa, B., Valvo, F.L., Ballesteros, G.A., Barberá, G.G., Amengual, J.F., Rodriguez, A., McMinn, M. (2010). Recapture processes and biological inference in monitoring burrow-nesting seabirds. Journal of Ornithology 151: 133-146.
Sanz-Aguilar, A., Mínguez, E., Oro, D. (2012) Is laying a large egg expensive? Female-biased cost of first reproduction in a petrel. The Auk, 129: 510-516.
Sanz-Aguilar, A., Marco Jover, E., Linares Molina, J., Grau, C., Izquierdo, A., Mascarell, J., Pellicer, A. (2018a). Seguimiento de las colonias reproductoras de Paíño europeo (Hydrobates pelagicus melitensis) en la isla de Benidorm (ZEPA E-121) P.N. Serra Gelada. Informe de actividades y resultados obtenidos en 2018. Dirección General de Gestión del Medio Natural. Generalitat Valenciana. Informe inédito.
Sanz-Aguilar, A., Payo-Payo, A., Igual, J.M., Rotger, A., Negre, N., Picorelli, V., Viñas, M., Martínez, O., Cardona, E., Luna, R. (2018b). Estudi de la influència de les paparres (Ornithodoros maritimus) sobre els paràmetres demogràfics dels fumarells (Hydrobates pelagicus) a la colònia de s’Espartar. Informe d’activitats i resultats de la campanya. Govern Balear. Informe inédito.
Scott, D. (1970) The breeding biology of the Storm Petrel Hydrobates pelagicus.Tesis doctoral. Universidad de Oxford, UK.
Sin-Yeon, K., Sanz-Aguilar, A., Mínguez, E., Oro, D. (2012). Small-scale spatial variation in evolvability for life-history traits in the storm petrel. Biological Journal of the Linnean Society, 106: 439-446.
Soldatini, C., Albores-Barajas, Y.V., Massa, B., Gimenez, O. (2014). Climate driven life histories: the case of the mediterranean storm petrel. PLOS ONE, 9: e94526.
Tavecchia, G., Mínguez, E., De León, A., Louzao, M., Oro, D. (2008). Living close, doing differently: small-scale asynchrony in demography of two species of seabirds. Ecology, 89:77-85.
Tenan S, Pradel R, Tavecchia G, Igual JM, Sanz-Aguilar A, Genovart M, Oro D. (2014). Hierarchical modelling of population growth rate from individual capture-recapture data. Methods in Ecology and Evolution, 5: 606-614.
Zabala, J., Zuberogoitia, I., Martínez-Climent, J.A., Etxezarreta, J. (2011). Do long lived seabirds reduce the negative effects of acute pollution on adult survival by skipping breeding? A study with European storm petrels (Hydrobates pelagicus) during the “Prestige” oil-spill. Marine Pollution Bulletin, 62: 109-115.
Zuberogoitia, I., Crespo, A., Burgos, G., Zabala, J., Etxezarreta, J., Zuberogoitia, J. (2014). El paíño europeo Hydrobates pelagicus en Aketx, Bermeo (Bizkaia). 26 años de seguimiento: 1989-2014. Diputación Foral de Bizkaia. Informe inédito.
Zuberogoitia, I., Zabala, J., Etxezarreta, J., Crespo, A., Burgos, G., Arizaga, J. (2016). Assessing the impact of extreme adverse weather on the biological traits of a European storm petrel colony. Population Ecology, 58: 303-313.
Ana Sanz-Aguilar, Iñigo Zuberogoitia, Ángel Sallent,
Virginia Picorelli, Juan G. Navedo y Rafa Garaita
UIB & IMEDEA CSIC-UIB; Estudios Medioambientales Icarus S.L.; ANSE; Reserves naturals des Vedrà, es Vedranell i els illots de Ponent; Aves Cantábricas S.L.
Fecha de publicación: 4-06-2019
Sanz-Aguilar, A., Zuberogoitia, I., Sallent, Á., Picorelli, V., Navedo, J.G., Garaita, R. (2019). Paíño europeo – Hydrobates pelagicus. En: Enciclopedia Virtual de los Vertebrados Españoles. López, P., Martín, J., González-Solís, J. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
