|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: Loggerhead, reproduction, demography.
Biología de la reproducción Los patrones reproductivos de las tortugas marinas están fuertemente condicionados y relacionados con las migraciones a las playas de puesta (Spotila et al., 1997). La tortuga boba no se suele reproducir todos los años y suele dejar 1 ó 2, raramente 3 temporadas de reposo y alimentación en sus áreas de migración normalmente alejadas miles de kilómetros de las zonas de anidación. Aunque no se conocen en detalle sus ciclos gonadales es probable que la vitelogénesis se inicie pronto durante esta migración reproductora. Se ha estimado una frecuencia media de anidación de alrededor de dos años, aunque parece que esta frecuencia varía con el tiempo tanto a nivel poblacional como individual en función de condiciones ambientales como la temperatura del mar o la abundancia de alimento. Sin embargo, es probable que los machos se apareen todos los años. Es frecuente observar a los machos en el mar cerca de las playas durante la anidación esperando a las hembras que vuelven a ovular pocas horas después de cada puesta. La reproducción de la tortuga boba es estacional y en el Atlántico norte y el Mediterráneo se produce la puesta a final de primavera y verano mientras que las eclosiones se inician en verano y se prolongan durante el otoño (Varo-Cruz et al., 2007). Los apareamientos se inician temprano en la primavera durante la migración reproductora y son muy frecuentes cerca de las playas de anidación y especialmente al principio de la temporada de puesta, aunque continúan durante toda la época de puesta. El cortejo y las interacciones sexuales son poco conocidos pero mediante estudios de multipaternidad se sabe que muchas hembras se aparean con varios machos (Sanz et al., 2006). Además, se ha constatado la existencia de almacenamiento de esperma pues sucesivos nidos de la misma hembra pueden compartir los mismos padres (Sanz et al., 2007). Muy probablemente tenga lugar competición espermática para fecundar los huevos dado que suele haber una clara dominancia en número de huevos fecundados por el esperma de uno de los padres de cada nido, dominancia que se suele mantener en los nidos sucesivos de una hembra dada (Sanz et al., 2007). La época de puesta es estival y suele durar unos 3 meses iniciándose en abril (Florida), mayo (Mediterráneo) o fin de junio (Cabo Verde). Durante este periodo cada hembra realizará entre 1 y 7 nidos con una media de 3 a 5 según zonas. En Chipre, las hembras más grandes ponen más huevos pero no tienen más nidos por temporada (Broderick et al., 2003). En el Mediterráneo parece que la frecuencia de puestas es menor que en Cabo Verde o Florida (Broderick et al., 2003; Varo-Cruz et al., 2007). Tras cada puesta se produce una nueva ovulación y fertilización de los huevos que pasan del ovario al oviducto donde adquirirán el albumen y formaran la cáscara durante los días que separan una puesta de la siguiente (Miller, 1997). El intervalo de tiempo entre anidaciones consecutivas puede variar entre 10 y 18 días. Temperaturas altas del agua pueden reducir sensiblemente el tiempo que transcurre entre dos puestas sucesivas (Hays et al., 2002).
Figura 1. Hembra desovando en una playa en Cabo Verde. (C) A. Marco
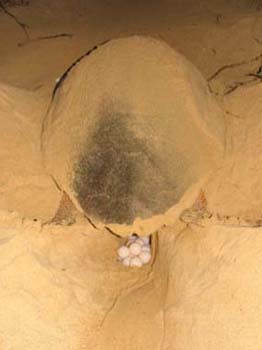
La anidación es nocturna y se puede ver afectada por las mareas, la luna o la iluminación artificial de la playa. Las hembras salen de forma aislada a la playa y ascienden lentamente seleccionando su lugar de puesta. Pueden realizar varios intentos de anidación antes de realizar el nido y también pueden retornar al agua sin anidar. El éxito de anidación en playas de Cabo Verde puede variar entre el 20% y el 70 % (Díaz-Merry y López-Jurado 2004; Varo-Cruz et al., 2007) en función de la calidad de la playa y las condiciones ambientales. En el Mediterráneo se han descrito éxitos de anidación mucho menores (Erk’akan, 1993). Todo el proceso de anidación suele durar algo más de 1 h distribuyendo de forma más o menos equitativa entre la excavación del nido, la ovoposición y el tapado y camuflado del nido (Díaz-Merry y López-Jurado, 2004). La excavación del nido se inicia con movimientos laterales de todo el cuerpo que entierran a la tortuga entre 10 y 25 cm, superando la zona de arena seca. Posteriormente inician un excavado con las patas traseras de un huevo en forma de bombilla de unos 25 a 30 cm donde depositarán los huevos. La profundidad total media del fondo del nido respecto a la superficie de la arena suele ser entre 35 y 60 cm mientras que la parte superior del nido suele estar entre 20 y 35 cm de la superficie. La liberación de los huevos suele ser de forma individual o en grupos de 2, 3 o incluso 4 huevos de golpe y suele durar unos 30 minutos. La liberación de cada grupo de huevos suele ir acompañada de contracciones abdominales de la hembra y de un fluido oviductal transparente que parece dar cierta protección inicial a los huevos. Tras la puesta tapan rápidamente los huevos con arena usando las patas traseras, compactan la arena con el peso de su cuerpo y el empuje de sus extremidades y camuflan el lugar del nido removiendo arena con las aletas delanteras durante un periodo medio de 20-25 min. Tras este proceso retornan al mar dejando sus rastros particulares en la arena caracterizados por su tamaño y el movimiento alterno de las extremidades de cada lado. El número de huevos de cada puesta es variable geográficamente de forma que en Florida suelen poner entre 100 y 125 huevos por nido mientras que en Cabo Verde o Grecia suelen poner entre 65 y 95 huevos. Las hembras americanas son también sensiblemente más grandes que las africanas o europeas. De forma excepcional hay hembras que pueden hacer nidos muy pequeños de hasta 30 huevos o muy grandes de hasta 180 (LeBuff, 1990). Se ha observado en diferentes zonas una variabilidad importante entre años en el tamaño medio de puesta que probablemente se deba a factores ambientales que influyen en la adquisición de alimento de las madres (LeBuff, 1990; Varo-Cruz et al., 2007). Los huevos son esféricos, blancos, levemente cremosos y apergaminados, con poca elasticidad. Tienen un diámetro medio de unos 40 mm (entre 35 y 55 mm ) y un peso inicial medio de entre 25 y 50 g (Dodd, 1988), dimensiones que variarán levemente durante la incubación. Es rara la formación y deposición de falsos huevos sin yema ni embrión como ocurre con la tortuga laúd. Al inicio se suele dar una cierta deshidratación de los huevos que se puede compensar posteriormente. La tasa de fertilización de los huevos suele ser muy alta en esta especie, superando con frecuencia el 90 % (Miller, 1997; Abella et al., 2007c). Tras las primeras horas en el nido, los embriones emigran hacia la parte superior del huevo donde se fijarán e iniciarán la formación de las membranas perivitelinas (Miller, 1997). Este proceso va acompañado de la formación en esa zona apical de una mancha blanca en la cáscara con un diámetro que irá aumentando durante los primeros días de incubación hasta cubrir todo el huevo hacia el día 10 (Abella et al., 2006). Si el huevo muere esta coloración blanca muy contrastada con el tono más cremoso de huevos recién puestos o muertos se difumina y desaparece. La incubación dura en término medio unos 52-56 días a unos 29,5 ºC de temperatura en los que se da una razón de sexos equilibrada (Mrosovsky, 1988) auqnue puede oscilar entre 46 y 70 días en función, sobre todo, de la temperatura de incubación. Temperaturas superiores aceleran el desarrollo y si son durante el segundo tercio de la incubación producen más hembras mientras que condiciones más frías retrasan el proceso y producen más machos. Por encima de 32 grados C o por debajo de 27 ºC se producirían individuos de un sólo sexo (Mrosovsky et al., 2002). La eclosión es un proceso lento que se inica con el picado y rasgado de la cáscara con el diente córneo provisional del rostro. El neonato irá saliendo poco a poco del huevo con movimientos separados con periodos de reposo que pueden sumar hasta la salida total del huevo más de dos días. Este proceso, junto con la emergencia vertical en grupo del nido hasta la superficie puede durar en media 4 días. Algunos neonatos emiten pitidos dentro del nido que podrían tener algún papel en la sincronización de la eclosión o la emergencia. La emergencia suele ser masiva y nocturna aunque algunas tortugas tardías pueden demorar su emergencia hasta varios días. Los neonatos tienen una longitud media del espaldar de entre 35 y 45 mm y un peso medio de entre 15 y 20 gr. El tamaño del huevo determina, en gran medida, la longitud y el peso de las crías que nacen, de forma que huevos más grandes producen crías más pesadas y más largas (Odzemir et al., 2007). Tras emerger se desplazan con sus patas sobre las dunas y la playa, sin pausa hasta el mar, al que pueden llegar en pocos minutos. Nidos alejados de la orilla o la presencia de basura, estructuras o iluminación artificial en la playa pueden alterar e impedir el acceso al agua y la provocar la muerte de las tortuguitas.
Estructura y dinámica de poblaciones El éxito de emergencia de crías es enormemente variable entre nidos, entre playas y estacionalmente, influyendo decisivamente factores como la depredación, la inundación, la erosión de la arena, la deshidratación excesiva del nido, la infección por microorganismos o la presencia de arcilla o raíces de plantas (Wallace et al., 2004, Marco et al., 2007; Abella et al., 2008). Se han detectado variaciones de < 20 a >75 % de éxito de eclosión entre playas contiguas en poblaciones de Cabo Verde y Mediterráneo (Margaritoulis, 2005; Varo-Cruz et al., 2007). La mortalidad de huevos y de neonatos en los primeros momentos de vida libre suele ser muy elevada pudiendo superar el 90%. Se estima que la probabilidad de retorno de un neonato a reproducirse puede ser en muchos casos menor del 1 por mil aunque esta baja supervivencia podría ser sostenible en condiciones naturales al ser una especie con una fertilidad (hasta más de 10.000 huevos totales por hembra) y longevidad (hasta más de 40 años fértiles) elevadas. Debido a la determinación fenotípica del sexo durante la incubación, la razón de sexos depende de la temperatura de incubación y suele estar sesgada hacia hembras en muchas zonas de anidación, encontrándose incluso playas donde no se producen machos (Godley et al., 2001; Abella et al., 2007a). Este hecho hace que el cambio climático se perfile como un gran riesgo para las tortugas marinas. Sin embargo, la razón de sexos de juveniles en algunas zonas de alimentación aparece casi equilibrado o incluso sesgado a machos (Casale et al., 2002). Una mortalidad mayor de hembras juveniles, una mayor capturabilidad de machos o una migración asimétrica de machos o hembras podrían explicar este fenómeno. Se ha estimado en el noroeste del Atlántico que duración mínima de la fase oceánica se extiende hasta los 7 años de edad y una longitud del caparazón de 46 cm. La duración del estadio nerítico es de 17 años en el noroeste del Atlántico, con una longitud del caparazón de 49-90 cm. Se ha estimado que la edad de madurez sexual en el noroeste del Atlántico varía entre 22 y 38,8 años y unos 92 cm de longitud del caparazón. En las poblaciones del Mediterráneo se estima una edad de la madurez de 19 a 30 años y 66,5 cm de longitud del caparazón (Casale et al., 2011b, Piovano et al., 2011; Avens y Snover, 20131). En Cabo Verde los tamaños corporales a la madurez son similares, pero un estudio de anillos de crecimiento en los huesos sugiere una maduración más temprana de algunos individuos con menos de 20 anillos óseos (Eder et al., 2012)1. El amplio rango de edad de la madurez se debe a los diferentes métodos de estima utilizados (marcaje-recaptura, esqueletocronología; Avens y Snover, 20131). Las importantes diferencias entre tamaño medio de los adultos reproductores entre América, África y Europa podrían responder a una diferente edad de maduración pero también podría ser debido a una diferente tasa de crecimiento o a ambos factores. La alta edad de madurez sexual y la baja supervivencia de huevos y crías recién nacidas, indican que la supervivencia de juveniles y adultos debe ser elevada para mantener a las poblaciones. Estudios de marcaje y recaptura estiman una mortalidad anual de juveniles en estas aguas del 27 % (Casale et al., 2007) en el Mediterráneo. En el Atlántico, para juveniles en vida oceánica la supervivencia anual podría superar el 85%, mientras que para juveniles de mayor tamaño con vida nerítica, la superviencia anual sería menor del 80 %. Se estima que si los juveniles de tortuga boba permanecen un tiempo medio de 15 años en aguas mediterráneas, tendrían una supervivencia global menor a un 1%. La supervivencia anual de hembras adultas en el Atlántico norte podría superar el 80 % (Heppell et al., 2003), mientras que la supervivencia de machos adultos sigue sin conocerse. En zonas de anidación donde se captura a las hembras en la playa, la supervivencia de adultos podría ser mucho menor. En Cabo Verde, se ha estimado una mortalidad anual de hembras anidantes superior al 35 % (Marco et al., 2008). Se ha estimado una tasa anual de reclutamiento de nuevas hembras adultas a las playas de anidación de entre un 10 y un 30 %, renovándose la mitad de la población de hembras cada 3 a 6 años. De estos datos se deduce que impactos ambientales están provocando un rejuvenecimiento de las poblaciones, de forma que la mayoría de las tortugas no llegue a los 30 años de vida. Sin embargo, se estima que la duración de la vida fértil potencial en hembras podría ser de unos 20 a 30 años que sumado a los 10 a 35 años estimados de vida juvenil suponen una longevidad de unos 45-50 años que podría llegar en casos extremos hasta los 70 años (Frazer, 1987; Dodd, 1988). Las altas tasas de mortalidad juvenil podrían ser insostenibles y provocar el colapso de la reproducción en algunas poblaciones. La alta mortalidad juvenil unida a la gran filopatría natal de la especie puede estar también dificultando enormemente la dispersión de la anidación y la colonización de nuevas playas reproductoras.
Referencias Abella, E., Marco, A., Diéguez-Uribeondo, J. López-Jurado, L. F. (2008). Pathogenic effect of microorganisms on loggerhead eggs. En: Abstracts of 28th Sea Turtle Symposium. International Sea Turtle Society, Loreto, Baja California Sur, Mexico. Abella, E., Marco, A., López-Jurado, L. F. (2006). Why are not correlated egg fertilization rate and hatching success in sea turtles? En: Abstracts of 26th Sea Turtle Symposium. International Sea Turtle Society. Island of Crete, Greece. Abella, E., Marco, A., López-Jurado, L. F. (2007a). Climate change and the evolution of loggerhead sex-ratio in Cabo Verde. Abstracts of 14th European Congress of Herpetology. Societas Europaea Herpetologica. Lisboa, Portugal. Abella, E., Marco, A., López Jurado, L. F. (2007c). Success of delayed translocation of loggerhead turtle nests: implications in management programs. J. Wild. Manag., 71: 2290-2296. Avens, L., Snover, M. L. (2013). Age and Age stimation in Sea Turtles. Pp. 97-133. En: Wyneken, J., Lohmann, K. J., Musick, J. A. (Eds.). The Biology of Sea Turtles. Volume III. CRC Press, Boca Raton. Broderick, A.C., Glen, F., Godley, B.J., Hays, G.C. (2003). Variation in reproductive output of marine turtles. J. Exp. Mar. Biol. Ecol., 288: 95-109. Casale, P., Laurent, L., Gerosa, G., Argano, R. (2002). Molecular evidence of male-biased dispersal in loggerhead turtle juveniles. J. Exp. Mar. Biol. Ecol., 267: 139-145. Casale, P., Mazaris, A. D., Freggi, D. (2011). Estimation of age at maturity of loggerhead sea turtles Caretta caretta in the Mediterranean using length-frequency data. Endangered Species Research, 13 (2): 123-129. Casale, P., Mazaris, A. D., Freggi, D., Basso, R., Argano, R. (2007). Survival probabilities of loggerhead sea turtles (Caretta caretta) estimated from capture-mark-recapture data in the Mediterranean Sea. Sci. Mar., 71: 365-372. Díaz-Merry, A., López-Jurado, L.F. (2004). Temporary sequences of ovoposition in loggerhead females from the Cape Verde islands. Pp. 249-251. Proceedings of the 21st Annual Symposium on Sea Turtle Biology and Conservation. NOAA Tech. Memo. NMFS-SEFSC-528. Philadelphia, Pennsylvania, U.S.A. Dodd, C. K. Jr. (1988). Synopsis of the biological data on the loggerhead sea turtle Caretta caretta (Linnaeus, 1758). U. S. Fish Wildl. Serv. Biol. Rep., 88 (14): 1-110. Eder, E., Ceballos, A., Martins, S., Pérez-García, H., Marín I, Marco, A., Cardona, L. (2012). Foraging dichotomy in loggerhead sea turtles Caretta caretta off northwestern Africa. Marine Ecology Progress Series, 470: 113-122. Erk’akan, F. (1993). Nesting biology of loggerhead turtles Caretta caretta L. on Dalyan beach, Mugla-Turkey. Biol. Conserv., 66: 1-4. Frazier, N. (1987). Preliminary estimates of survivorship for wild loggerhead sea turtles (Caretta caretta). J. Herpetol., 21: 232-235. Godley, B. J., Broderick, A. C., Mrosovsky, N. (2001). Estimating hatchling sex ratios of loggerhead turtles in Cyprus from incubation durations. Mar. Ecol. Progr. Ser., 210: 195–201. Hays, G. C., Broderick, A. C., Glen, F., Godley, B. J., Houghton, J. D. R., Metcalfe, J. D. (2002). Water temperature and internesting intervals for loggerhead (Caretta caretta) and green (Chelonia mydas) sea turtles. J. Thermal Biol., 27: 429-432. Heppell, S., Snover, M. L., Crowder, L. B. (2003). Sea turtle Popoulation Dynamics. Pp. 275-306. En: Lutz, P. L., Musick, J. A., Wyneken, J. (Eds.). The Biology of Sea Turtles. Volume II. CRC Press Boca Raton, Florida, USA. Le Buff, C. R. Jr. (1990). The loggerhead turtle in the Eastern Golf of Mexico. Caretta Research, Sanibel Florida, USA. 216 pp. Marco, A., Abella, E., López-Jurado, L. F. (2007). Vulnerability of turtle eggs to the presence of clay in nesting beaches. Proceedings of the 27th International Symposium on Sea Turtles. International Sea Turtle Society. Myrtle Beach, Carolina, USA. Marco, A., López, O., Abella, E., Varo, N., Martins, S., Gaona, P., Sanz, P., López-Jurado, L.P. (2008). Massive capture of nesting females is severely threatening the caboverdian loggerhead population. Proceedings of the 28th International Symposium on Sea Turtles. International Sea Turtle Society. Loreto, Baja California Sur, Mexico. Miller, J.D. (1997). Reproduction in sea turtles. Pp. 51-81. En: Lutz, P.L., Musick, J.A. (Eds.). The Biology of sea turtles. CRC Press Boca Ratón, Florida, USA. Mrosovsky, N. (1988). Pivotal temperatures for loggerhead turtles Caretta caretta from northern and southern nesting beaches. Can. J. Zool., 66: 661–669. Mrosovsky, N., Kamel, S., Rees, A. F., Margaritoulis, D. (2002). Pivotal temperature for loggerhead turtles (Caretta caretta) from Kyparissia Bay, Greece. Can. J. Zool., 80: 2118-2124. Odzemir, A., Ilgaz, C., Kumlutas, Y., Durmus, S. H., Kaska, Y., and Turkozan, O. (2007). An assessment of initial body size in loggerhead sea turtle (Caretta caretta) hatchlings in Turkey . Zool. Sci., 24: 376-380. Piovano, S., Clusa, M., Carreras, C., Giacoma, C., Pascual, M., Cardona, L. (2011). Different growth rates between loggerhead sea turtles (Caretta caretta) of Mediterranean and Atlantic origin in the Mediterranean Sea. Marine Biology (Berlin), 158 (11): 2577-2587. Sanz, P., Roques, S., Marco, A., López-Jurado, L. (2006). Estudio genético de paternidad múltiple de la tortuga común Caretta caretta en el Archipiélago de Cabo Verde: aplicación a la ecología de la conservación. Actas del IX Congreso Luso-Español y XIII Congreso Español de Herpetología. Asociación Española de Herpetología, San Sebastián, España. Sanz, P., Roques, S., Marco, A., López-Jurado, L.F. (2007). Fine-scale paternity study of a loggerhead from Cape Vert: within and between seasons. Proceedings of the 27th International Symposium of Sea Turtles. International Sea Turtle Society. Myrtle Beach, Carolina, USA . Spotila, J., O’ Connor, M. J., Paladino, F. V. (1997). Thermal Biology. Pp. 299-316. En: Lutz, P. L., Musick, J. A. (Eds.). The Biology of Sea Turtles. CRC Press Boca Ratón, Florida, USA. Varo-Cruz, N., Cejudo, D., López-Jurado, L. F. (2007). Reproductive biology of the loggerhead turtle (Caretta caretta L. 1758) on the island of Boavista (Cape Verde, West Africa). Pp. 127-144. En: López-Jurado, L. F., Liria-Loza, A. (Eds.). Marine Turtles. Recovery of Extinct Populations. Monografías del Instituto Canario de Ciencias Marinas, 5. Las Palmas, España. Wallace, B. P., Sotherland, P.R., Spotila, J. R., Reina, R.D., Franks, B. F., Paladino F. V. (2004). Abiotic and biotic factors affect the nest environment of embryonic leatherback turtles, Dermochelys coriacea. Physiol. Biochem. Zool., 77: 423-432.
Adolfo Marco, Carlos Carreras, Elena Abella Fecha de publicación: 24-06-2008 Otras contribuciones: 1. Alfredo Salvador. 16-11-2015 Marco, A., Carreras, C., Abella, E. (2015). Tortuga boba – Caretta caretta. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Marco, A. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|
|
|
|
|
|