|
|
||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Spur-thighed tortoise, description, measurements, mass, variation.
Descripción del adultoEs un quelonio de caparazón abombado cuyo espaldar es de color amarillento, verde oliva pálido o pardusco, en el que las placas presentan generalmente una mancha central y los bordes de color oscuro o negro. El peto es amarillento, con manchas irregulares negras, que en algunos individuos pueden extenderse ampliamente alrededor de las suturas centrales, mientras que en otros forman un diseño estrellado especialmente en las placas ventrales y femorales.
Figura 1. Distintos diseños dorsales y ventrales de Testudo graeca de Doñana.© A.C. Andreu, y A. Portheault.
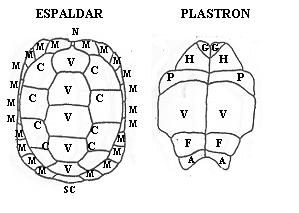
En el espaldar hay cinco placas vertebrales, ocho costales en dos series de cuatro a ambos lados de las vertebrales, once marginales al lado de cada serie costal, una nucal y una supracaudal. Ésta última no está divididida ni sobresale de las marginales. En el plastron se distinguen dos placas gulares, dos humerales, dos pectorales, dos ventrales, dos femorales y dos anales. Las escamas ventrales son las de mayor tamaño. La parte ósea posterior del peto, el xiphiplastron, presenta cierta movilidad gracias a una charnela situada entre las placas femorales y las abdominales. La cabeza es de color amarillento, con manchas negras o completamente negra, y se encuentra cubierta por pequeñas e irregulares escamas, entre las que destacan la frontal por su mayor anchura (a veces dividida) y la prefrontal. Las supranasales no se encuentran en la zona media. Las mandíbulas, con un recubrimiento córneo, conforman una boca en forma de pico, sin dientes.
Figura 2. Placas que componen el espaldar y plastron de la tortuga mora. En el espaldar: N: nucal, M: marginales, V: vertebrales, C: costales, SC: supracaudal. En el plastron: G: gulares, H: humerales, P: pectorales, V: ventrales, F: femorales, A: anales.
Figura 3. Detalles de la cabeza, cola, espolón del muslo, escamas imbricadas y uñas de miembro anterior, y del miembro posterior de una tortuga mora de Doñana. © A. Portheault.
Las patas están cubiertas de pequeñas escamas, aunque las delanteras presentan en su parte anterior tres a seis filas de grandes escamas puntiagudas e imbricadas. Un tubérculo córneo, subcónico y a veces de aspecto aplanado por el desgaste, en la parte trasera de los muslos es característico de esta especie. Posee cinco uñas en las patas delanteras y cuatro en las traseras. La cola no presenta uña terminal (Andreu y López-Jurado, 1997).
Tamaño corporal En las poblaciones ibéricas, los individuos no suelen sobrepasar los 200 mm de longitud de espaldar. En Doñana, los machos tienen un tamaño medio de 146mm y peso medio de 558 g, mientras que las hembras suelen tener mayor tamaño, promediando 173 mm de longitud de espaldar y 866 g de peso (Andreu, 1987; Díaz-Paniagua, Keller y Andreu, datos no publicados) (Díaz-Paniagua y Andreu, 2009). En la población del sureste ibérico, el tamaño corporal medio de los adultos de la Galera (Murcia) es de 113 mm para los machos y 134 para las hembras, no superando los 170mm (A. Giménez, datos no publicados) (Díaz-Paniagua y Andreu, 2009). En Marruecos, El Mouden et al. (2002) describen el tamaño del caparazón de los machos adultos entre 100,8 y 156,0 mm, con masa corporal entre 215 y 880 g; en las hembras el caparazón varía entre 100,5 y 186,0 mm y su masa corporal entre 201 y 1.337 g.
Tabla 1. Descripción general del tamaño de las tortugas adultas de Doñana (las longitudes de Doñana corresponden a datos de Andreu,1987; los pesos se han promediado de datos de campo obtenidos entre 1985 y 1999), Murcia (datos no publicados de la Reserva de la Galera cedidos por A. Giménez), y de Marruecos (El Mouden et al., 2002). Se presenta la media + desviación típica y valor mínimo y máximo, excepto en las de Murcia donde no se da la desviación típica.
Dimorfismo sexualLas hembras alcanzan en general mayor tamaño corporal que los machos, pues la edad de madurez es más tardía y presentan por tanto mayor periodo de crecimiento juvenil. Los machos adultos se diferencian fácilmente de las hembras por la forma de las escamas anales, que tienen menor longitud que en las hembras, y frecuentemente presentan un aspecto desgastado en el borde más próximo a la cola. En las hembras, estas escamas son más alargadas y puntiagudas, llegando casi a tocar el espaldar. Las suturas entre las placas femorales y anales son aproximadamente rectas en los machos, mientras que en las hembras forman un ángulo dirigido hacia la parte anterior del cuerpo. El peto de los machos es más cóncavo y la placa supracaudal convexa, fuertemente curvada hacia dentro, mientras que en las hembras tanto el peto como la supracaudal son planos (Angel, 1946). Braza et al. (1981) hallaron que el crecimiento de la cabeza es diferente en hembras y machos, presentando finalmente éstas la cabeza más corta y ancha, si bien este carácter no es fácil de distinguir a simple vista.
Figura 4. Detalle de las escamas anales y femorales de una tortuga macho (A) y de una hembra (B).C) La escama supracaudal de los machos está fuertemente curvada hacia dentro.© A. Portheault.
Descripción de las crías Las crías nacen con un tamaño medio de 34,14mm (sd =1,92, rango: 28,7-39,9, n=26) y una masa corporal media de 10,8 g (sd = 2,01, rango 8,1-15,6mm, n=26) (Díaz-Paniagua et al., 1997). Tras la eclosión se aprecia un surco transversal arrugado en la zona media del peto, que recuerda la posición doblada que el animal ha tenido durante su crecimiento en el interior del huevo. La coloración del espaldar es similar a la del adulto, aunque las manchas oscuras del peto no se encuentran todavía completamente desarrolladas, variando entre algunos individuos que presentan escasas manchas y otros que muestran una irregular mancha situada alrededor del ombligo. En las crías recién nacidas se aprecian restos del surco umbilical y una protuberancia angulosa en el pico, que se va perdiendo aproximadamente en el primer o segundo mes de vida (datos propios) (Díaz-Paniagua y Andreu, 2009).
Figura 5. © A. Conejo.
Figura 6. Distintos diseños ventrales de las crías recién nacidas de tortuga mora. © C. Díaz-Paniagua.
Descripción de los huevosLos huevos son de color blanco y de forma aproximadamente esférica. Su diámetro medio mayor es de 33,7mm y el perpendicular a éste de 28,3 mm. El peso medio es de 14g (Díaz-Paniagua et al., 1996). (Ver Biología de la reproducción).La cáscara de los huevos de la tortuga mora está formada por una cubierta calcárea rígida y mínimamente porosa. A través de estos poros se produce el intercambio de gases y agua con el exterior. Sin embargo, la cantidad de agua que estos huevos rígidos pueden absorber es muy pequeña o imperceptible (Packard y Packard, 1980), no incrementándose apenas el tamaño ni el peso del huevo a lo largo del desarrollo embrionario (datos propios no publicados). La microestructura de la cáscara del huevo de T. graeca ha sido descrita por Young (1950) y Schleich y Kästle (1988) y es en general muy parecida a los de otras especies de la familia Testudinidae. Desde el exterior hacía el interior se distinguen varios estratos o capas: 1) Una cutícula externa o tegumento, en la que se pueden apreciar poros; b) Un estrato cristalino compuesto por cristales de carbonato cálcico bajo la forma de aragonita (estos cristales están formados por un núcleo esférico basal del que irradian finas agujas o espículas); c) Una membrana testácea ; d) dos membranas internas.
Características genéticasLa descripción del cariotipo de Testudo graeca fue publicada por Shindarov et al. en 1976, a partir de ejemplares de Bulgaria, que actualmente se consideran una especie distinta, Testudo ibera (Obst y Ambrosius, 1971; Bour, 1988; Gmira, 1993; David, 1994). Es muy probable que el cariotipo descrito para estas tortugas (número cromosómico 2n=52: 20 cromosomas metacéntricos y submetacéntricos, 10 subtelocéntricos y 22 telocéntricos y acrocéntricos), sea muy similar al de T. graeca, ya que hasta el momento todas las especies estudiadas dentro de la familia Testudinidae poseen el mismo número cromosómico (n=52), con 9 pares de macrocromosomas metacéntricos o submetacéntricos, 5 pares de macrocromosomas telocéntricos o subtelocénticos y 12 pares de microcromosomas (Bickham y Carr, 1983). Las diferencias entre las descripciones de algunos trabajos se debe a una distinta interpretación en la asignación de unos cromosomas a uno u otro tipo. En ningún caso se han identificado cromosomas sexuales.
Variación geográfica Hasta hace pocos años, se pensaba que la especie, T. graeca, se extendía por un amplia área de distribución que incluía el norte de África, los países ribereños del Mediterráneo oriental, parte de la Europa oriental, Asia Menor y Central. Sin embargo, en los últimos años del siglo XX y primeros del XXI, un buen número de trabajos morfológicos y genéticos (Obst y Ambrosius, 1971; Bour, 1988; Gmira, 1993; Perälä, 2002), pusieron de manifiesto la naturaleza polifilética de T. graeca y por tanto la necesidad de elevar a especie a varias de las hasta entonces consideradas subespecies de T. graeca. En este contexto se ha considerado que las tortugas del noroeste de África y las españolas se incluyen en una especie distinta de las del Mediterráneo oriental, Asia Menor y Central, habiendo heredado la denominación específica de Testudo graeca (Obst y Ambrosius, 1971; Bour, 1988; Gmira, 1993; David, 2004). Dentro del área de distribución de la especie, varios autores han descrito diferentes formas asignándoles nivel subespecífico, específico e incluso genérico (Highfield y Martin, 1989a y b; Highfield, 1990; Ballasina y Van der Kuyl., 2001; Perälä, 2002; Pieh, 2000, 2002; Pieh y Perälä, 2002). La especie muestra una alta variabilidad morfológica lo que ha dado lugar a la descripción de hasta 20 tipos distintos y a la consiguiente descripción de diferentes subespecies, especies o incluso géneros (Pieh y Perälä, 2002; Fritz et al., 2007). Esta variabilidad parece responder a una elevada plasticidad fenotípica que se habría producido como respuesta adaptativa, a escala local, a diferentes condiciones ambientales. A esta conclusión se ha llegado tras recientes estudios de la variación genética de un amplio número de poblaciones a lo largo de todo el área de distribución, los cuales indican que todas las poblaciones existentes entre Marruecos e Irán pertenecen a la una sola especie politípica, Testudo graeca (Fritz et al., 2007, 2009; Mikulíček et al., 2013)2. Las poblaciones españolas de tortuga mora han sido habitualmente asignadas a la subespecie Testudo graeca graeca, junto a las de Marruecos, Túnez, Argelia y Libia (Loveridge y Williams, 1957; Wermuth, 1958; Wermuth y Mertens, 1961; Harris et al., 2003; Andreu et al., 2004). En estudios genéticos realizados sobre ADN mitocondrial de ejemplares procedentes de las poblaciones ibéricas y del norte de África, se han encontrado hasta nueve haplotipos distintos, muy cercanos entre sí y con una distribución que no concuerda con las subespecies morfológicas descritas (Álvarez et al., 2000; Ballasina et al., 2001; Van der Kuyl, 2002; Harris et al., 2003). En T. graeca hay seis clados que se diferenciaron hace unos 1,8 – 4,2 millones de años. Uno de ellos corresponde a las poblaciones del norte de África, que se diferenciaron en cuatro linajes hace unos 1,1 – 1,4 millones de años según periodos de aridez durante el Pleistoceno (Fritz et al., 2009).1 Las poblaciones del sureste peninsular se caracterizan por una disminución de la diversidad genética con la distancia al área de colonización y una fuerte diferenciación espacial (Graciá et al., 2013a)2. Actualmente las poblaciones ibéricas, las de Mallorca, la mayor parte de Argelia y zonas cercanas a Argelia del este de Marruecos, todas ellas con el mismo linaje de ADN mitocondrial (B1), se consideran incluidas en la subespecie Testudo graeca graeca (Fritz et al., 2009). Esta subespecie se caracteriza por ser de talla mediana y color amarillento, oliva pálido o pardo. Las escamas del espaldar tienen, por lo general, sus bordes negros u oscuros y una mancha central también negra; las escamas del peto presentan manchas negras irregulares. A su vez, estas poblaciones forman parte de un grupo más amplio, las tortugas moras del Mediterráneo occidental, aisladas geográficamente del resto de poblaciones de T. graeca por el desierto de Libia y que, según indican los análisis de ADN nuclear y mitocondrial, forman un clado claramente delimitado dentro de la especie (Fritz et al., 2007, 2009; Mikulíček et al., 2013). Dentro de este grupo, además de la subespecie nominal, se reconocen actualmente cuatro subespecies más: T. g. nabeulensis Pieh y Perälä, 2004, presente en el este de Argelía, Tunez, Cerdeña y Sicilia; T. g. cyrenaica Pieh y Perälä, 2002, aislada en la península de Cirenaica, en Libia noroccidental; T. g. soussensis Pieh, 2000, del sur de Marruecos y T. g. marokkensis Pieh y Perälä, 2004, del noroeste de Marruecos (Fritz et al., 2009)2. Origen de las poblaciones españolasEl origen de las poblaciones españolas de tortugas moras fue objeto de controversia durante buena parte del siglo XX. La distribución muy restringida de estas poblaciones en la Península Ibérica (son las únicas poblaciones continentales de la especie en Europa) y su similitud morfológica con las tortugas del norte de Marruecos, sus conespecíficas más cercanas, indicaban una estrecha relación filogenética con éstas. Se propusieron varias hipótesis. Por un lado, se sugería que las poblaciones ibéricas serían los últimos relictos de una más extendida y amplia distribución de la especie en tiempos anteriores en la península. El origen de la especie habría sido Eurasia, habiendo quedado las poblaciones ibéricas en algún momento desconectadas de las de Oriente. Estas poblaciones habrían colonizado desde la península el norte de África (Bons, 1967; Loveridge y Williams, 1957). Otra hipótesis abogaba por un origen africano de las poblaciones ibéricas (Boscá, 1877; Lortet, 1887; Salvador, 1974). Otra cuestión por resolver se refería al momento de la separación entre las poblaciones españolas y africanas. Existían dos posibilidades. La primera de ellas consideraba que las poblaciones ibéricas y las norteafricanas se habrían separado antes de la apertura del estrecho de Gibraltar, suceso que tuvo lugar hace algo más de 5 millones de años, lo cual habría dado lugar a algún tipo de diferenciación genética y/o morfológica entre las tortugas de uno u otro lado. La otra posibilidad es que tuvieran un origen común y próximo en el tiempo, es decir, las poblaciones de un lado del estrecho habrían dado origen a las del otro, en algún momento posterior a la apertura del istmo, el cual habrían atravesado bien cruzando de forma natural, a la deriva sobre vegetación flotante o incluso flotando sobre el agua directamente, bien de la mano del hombre. Si esto fuera cierto sería de esperar una completa identidad tanto genética como morfológica entre las tortugas de ambos lados del estrecho. En la revisión del registro fósil pleistocénico, no se ha encontrado ningún resto de T. graeca en la Península Ibérica, siendo abundantes sin embargo en Marruecos (Gmira, 1995). En comparación, la otra especie de tortuga terrestre peninsular, la tortuga mediterránea, T. hermanni, cuenta con un abundante y extendido registro fósil en España. A este respecto es conveniente aclarar que en 1986 se publicó el hallazgo de un fósil de T. graeca encontrado en el yacimiento del Pleistoceno superior de Cueva Horá, Darro, Granada (Bailón, 1986). Este resto fue posteriormente reclasificado como T. cf. hermanni por el mismo autor (Bailón, 2001). La falta de fósiles ibéricos es un indicio claro a favor de la hipótesis de un origen extraeuropeo y relativamente reciente de las tortugas moras ibéricas. Álvarez et al. (2000) realizaron estudios genéticos que pretendían aclarar el origen de las poblaciones ibéricas de tortugas moras. Estos autores compararon varias secuencias de nucleótidos de ADN mitocondrial, en individuos de las dos poblaciones ibéricas y en otros procedentes de cuatro puntos distintos del norte de Marruecos. Los resultados mostraron que todos los ejemplares ibéricos compartían un mismo haplotipo, que también se encontraba en el NE de Marruecos. Además de éste, en Marruecos se encontraron tres haplotipos más. La falta de variación genética entre las tortugas de uno y otro lado del estrecho indica que el aislamiento entre ellas databa de un tiempo posterior a la apertura del estrecho, y por lo tanto que en algún momento, posterior a este evento, las tortugas cruzaron el estrecho a través del mar. Por otra parte, el hecho de que las tortugas ibéricas, cuenten sólo con dos poblaciones y con un número relativamente pequeño de efectivos y no presenten diferencias ni al nivel genético estudiado ni en el morfológico, mientras que, en el norte de África, se da una mayor diversidad genética y morfológica y las poblaciones son más abundantes y extendidas, indica que las tortugas españolas provienen del norte de África y concretamente del NE de Marruecos, y que llegaron a la península en un pasado relativamente reciente (no superior a algunos miles de años), bien de forma natural, por deriva a través del mar, como en el caso de otras tortugas que han colonizado islas, bien traídas por el hombre (Álvarez et al., 2000). Las poblaciones españolas tienen un origen reciente a partir de las poblaciones del este de Marruecos y de Argelia, bien sea por introducción humana o por dispersión a través del mar en tiempos históricos o prehistóricos (Fritz et al., 2009).1 Por otra parte, las tortugas moras de Baleares, tampoco han sido encontradas en el registro fósil de las islas, lo cual sugiere también que llegaron en un tiempo relativamente reciente y probablemente, por cercanía, del norte de África. Así pues, las tortugas moras españolas proceden del norte de África, de donde llegaron probablemente hace algunos cientos o quizás miles de años. En la actualidad se encuentran completamente integradas en los ecosistemas en los que habitan. Recientemente Graciá et al. (2011, 2013a, 2013b) aportan evidencias a favor de una llegada natural de las tortugas del sureste ibérico a finales del Pleistoceno, procedentes de zonas costeras del oeste de Argelia.2
Referencias Alvarez, Y., Mateo, J. A., Andreu, A. C., Díaz-Paniagua, C. Díez, A., Bautista, J.M. (2000). Mithocondrial DNA Haplotyping of Testudo graeca on both continental sides of the Straits of Gibraltar. Journal of Heredity, 91: 39-41. Andreu A.C. 1987. Ecología y dinámica poblacional de la tortuga mora, Testudo graeca, en Doñana. Tesis Doctoral. Univ. Sevilla Andreu, A. C., Lopez-Jurado, L. F. (1997). Testudo graeca. Pp. 110-117. En Salvador, A. (Coord.) Reptiles. Fauna Ibérica, vol 10. Ramos, M. A. Et al. (Eds.)Museo Nacional Ciencias Naturales, CSIC. Madrid. Andreu, A.C., Díaz-Paniagua, C., Keller, C., Slimani, T., El Mouden, H. (2004). Testudo [graeca] graeca.- Manouria 7 (22), 17-18 Angel, F. (1946). Reptiles et Amphibiens. Faune de France, vol. 45. Pierre André Imprimerie. Librairie de la Faculté des Sciences. Fédération Française des Sociétés de Sciences Naturelles, Office Central de Faunistique. Paris. Bailón, S. (1986). Los anfibios y reptiles del yacimiento de Cueva Horá (Darro, Granada). Antropología y paleoeología humana, 4:131-155. Bailón, S. (2001). Revisión de la asignación a Testudo cf. graeca del yacimiento del Pleistoceno superior de Cueva Horá (Darro, España). Revista Española Herpetología, 15: 61-65. Ballasina, D., Van der Kuyl, A. C. (2001). What is a Testudo graeca?. Chelonii, 3: 130-132. Ballasina, D., Van der Kuyl, A. C., Dekker, J. T., Maas, J., Willemsen, R. E., Goudsmit, J. (2001). Phylogenetic relationships among the species of the genus Testudo (Testudines: Testudinidae) inferred from mitochondrial 12S rRNA gene sequences. Chelonii, 3: 117-129. Bickham, J. W., Car, J. L. (1983). Taxonomy and phylogeny of the higher categories of cryptodiran turtles based on a cladistic análisis of chromosomal data. Copeia, 1983: 918-932. Bons, J. (1967). Recherches sur le biogeographie et la biologie des amphibiens et des reptiles du Maroc (Tesis Doctoral). Montpellier, France: Universidad de Montpellier. Boscá, E. (1877). Catálogo de los anfibios y reptiles observados en España, Portugal, Islas Baleares. An. Soc. Esp.Hist. Nat. Madrid, 6: 39-67. Bour, R. (1988).Caractères diagnostiques offerts par le crâne des tortues terrestres du genre Testudo. Mesogée, 48:13-19. Braza , F. Delibes, M., Castroviejo, J. (1981). Estudio biométrico y biológico de la tortuga mora (Testudo graeca) en la Reserva Biológica de Doñana, Huelva. Doñana, Acta Vertebrata, 8: 15-41. Buskirk, J. R., Keller, C., Andreu, A. (2001). Testudo graeca Linnaeus, 1758 – Maurische Landschildkröte. Pp. 125-177. En : Fritz, U. (Ed.). Handbuch der Reptilien und Amphibien Europas. Band 3/IIIA. Schildkröten (Testudines). I. (Bataguridae, Testudinidae, Emydidae). Aula-Verlag, Wiebelsheim. David, P. (1994). Liste des reptiles actuels du monde. I. Chelonii. Dumerilia, I: 7-127. Díaz-Paniagua, C., Andreu, A. C. (2009). Tortuga mora – Testudo graeca. En: Enciclopedia Virtual de los Vertebrados Españoles. Carrascal, L. M., Salvador, A. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/ Díaz-Paniagua, C., Keller, C., Andreu, A. C. (1996).Clutch frequency, egg and clutch characteristics, and nesting activity of spur-thigheed tortoises, Testudo graeca, in southwestern Spain. Canadian Journal of Zoology, 74: 560-564. Díaz-Paniagua, C., C. Keller, Andreu, A. C. (1997). Hatching success, delay of emergence and hatching biometry of Testudo graeca in southwestern Spain. Journal of Zoology, London, 243: 543-553. El Mouden, H., Slimani, T., Ben Kaddour, K. (2002) Croissance et dimorphisme sexuel chez la tortue mauresque (Testudo graeca graeca L. 1758). Chelonii, 3: 325-330. Fritz, U., Harris, D. J., Fahd, S., Rouag, R., Gracia Martínez, E., Giménez Casalduero, A., Siroky, P., Kalboussi, M., Jdeidi, T. B., Hundsdoerfer, A. K. (2009). Mitochondrial phylogeography of Testudo graeca in the Western Mediterranean: Old complex divergence in North Africa and recent arrival in Europe . Amphibia-Reptilia, 30 (1): 63-80. Fritz, U., Hundsdörfer, A.K., Široký, P., Auer, M., Kami, H., Lehman, J., Mazanaeva, L.F., Türkozan, O., Wink, M. (2007). Phenotipic plasticity leads to incongruence between morphology-based taxonomy and genetic differentiation in western Palaearctic tortoises (Testudo graeca complex, Testudines, Testudinidae). Amphibia-Reptilia, 28 (1): 97-121. Gmira, S. (1993). Nouvelles données sur les espèces actuelles de Testudo (Chelonii, Testudinidae). Bull. Soc. Herp. Fr., 65-66: 49-56. Gmira, S. (1995). Étude des Chéloniens fossiles du Maroc. Cahiers de Paléonthologie. CNRS Editions, Paris. Graciá, E., Botella, F., Anadón, J. D., Edelaar, P., Harris, D. H., Giménez, A. (2013a). Surfing in tortoises? Empirical signs of genetic structuring owing to range expansion. Biology Letters, 9 (3): 2012-1091. Graciá, E., Giménez, A., Anadón, J.D., Botella, F. (2011). Genetic patterns of a range expansion: the spur-thighed tortoise Testudo graeca graeca in Southeastern Spain. Amphibia-Reptilia, 32(1): 49-61. Graciá, E., Giménez, A., Anadón, J.D., Harris, J.D., Fritz, U., Botella, F. (2013b). The uncertainty of Late Pleistocene range expansions in the western Mediterranean: a case study of the colonization of south-eastern Spain by the spur-thighed tortoise, Testudo graeca. Journal of Biogeography, 40 (2): 323-334. Harris, D. J., Znari, M., Mace, J.-C., Carretero, M. A. (2003). Genetic variation in Testudo graeca from Morocco estimated using 12S rRNA sequencing. Revista Española Herpetología, 17:5-9. Highfield, A. C.(1990). Tortoises of north Africa; taxonomy, nomenclature, phylogeny and evolution with notes on field studies in Tunisia. J. Chel. Herp., 1: 1-56. Highfield, A. C., Martin, J. (1989a). A revision of the Testudines of North Africa, Asia and Europe. Genus: Testudo. J. Chel. Herp., 1: 1-12. Highfield, A. C., Martin, J. (1989b). Testudo whitei Bennett 1836. New light on an old carapace - Gilbert White's Selborne tortoise re-discovered. J. Chel. Herp., 1:13-22. Lortet, L. (1887). Observations sur les tortues terestres et paludines du Bassin de la Méditerranée. Arch. Mus. Hist. Nat. Lyon, 4: 1-26. Loveridge, A., Williams, E. E. (1957). Revision of the African Tortoises and Turtles of the Suborder Cryptodira. Bull. Mus. of Comp. Zool. 115: 163-557. Mikulíček, P., Jandzik, D., Fritz, U., Schneider, C., Široký, P. (2013). AFLP analysis shows high incongruence between genetic differentiation and morphology-based taxonomy in a widely distributed tortoise. Biological Journal of the Linnean Society, 108 (1): 151-160. Obst, F. J., Ambrosius, H. (1971). Taxonomische Studien an europeischen Landschildkröten (Reptilia: Testudinidae) mit serologisch-immunologischen Methoden. Zool. Abh. Mus. Tierk. Dresden, 30: 297-331. Packard, G.C., Packard, M.J. (1980). Evolution of cleidoic egg among reptilian antecedents of birds. Amer. Zool., 20: 351-362. Perälä, J. (2002). The genus Testudo (Testudines: Testudinidae): phylogenetic inferences. Chelonii, 3: 32-39. Pieh, A. (2000). Testudo graeca soussensis,eine neue Unterart der Maurischen Landschildkrote aus dem Sousstal (Sudwest Marokko). Salamandra, 36: 209-222. Pieh, A. (2002). Remarks on the geographic and evolutionary variability of the Spurtighed tortoise (Testudo graeca) in northern Africa. Chelonii, 3: 109-116. Pieh, A. , Perälä, J. (2002). Variabilität von Testudo graeca Linnaeus, 1758 im Östlichen Nordafrika mit Beschreibung eines neuen Taxons von der Cyrenaika (Nordostlibyen).Herpetozoa, 15:328. Salvador, A. (1974). Guía de los anfibios y reptiles españoles. Icona, Madrid. Schleich, H. H., Kästle, W. (1988). Reptile eggshells. SEM Atlas. Gustav Fischer Verlag. Stuttgart. New York. Schleich, H.H., Kästle, W., Kabisch, K. (1996). Amphibians and reptiles of North Africa. Koeltz Scientific Books, Koenigstein. Shindarov, L., Todorov, S. L., Vasileva, V., Georgieva, M. (1976). Cytogenetic investigation of cells from primary kidney culture of Testudo graeca. Comptes Rendus de l’Academica Bulgare des Sciences, 29: 1695-1697. Van der Kuyl, A. C., Ballasina, D. L., Dekker, J. T., Maas, J., Willemsen, R. E., Goudsmit, J. (2002). Phylogenetic relationships among the species of the genus Testudo (Testudines: Testudinidae) inferred from mitochondrial 12D rRNA gene sequences. Molecular Phylogenetics and Evolution, 22: 174-183. Wermuth, H. (1958). Status und Nomenklatur der Maurischen Landschildkrote, Testudo graeca, in SW Asien und NO Afrika. Senck. Biol., 39: 149-153. Wermuth, H., Mertens, R. (1961). Schildkröten, Krokodile, Brückenechsen. Gustav Fischer Verlag, Jena. Young, J. D. (1950). The structure and some physical properties of the Testudinian eggshell. Proc. Zool. Soc. Lond., 120: 455-473.
Carmen Díaz-Paniagua y Ana C. Andreu Fecha de publicación: 29-07-2005 Otras contribuciones: 1. Alfredo Salvador. 21-07-2009; 2. Alfredo Salvador. 15-09-2015 Díaz-Paniagua, C., Andreu, A. C. (2015). Tortuga mora – Testudo graeca. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Marco, A. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||