|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Geoffroy’s bat, sexual dimorphism, size, geographic variation.
Nomenclatura La especie fue descrita por Etienne Geoffroy Saint-Hilaire a principios del siglo XIX. Se añade la inicial en el nombre científico de esta especie para distinguir a Etienne de su hijo Isidore, a quien debemos, entre otras, la descripción de Tadarida brasiliensis. (I. Geoffroy, 1824). Cuando E. Geoffroy describió Myotis emarginatus en 1806 era el primer Myotis pequeño que se conocía en la fauna europea (Tupinier, 2001). Curiosamente ha sido el último Myotis pequeño incorporado a la lista de fauna española (sin contar con los recientes descubrimientos moleculares de especies crípticas) ya en los 60. En 1963 Balcells encontró dos colonias de cría en Huesca y Lérida, datos que salen publicados en Balcells (1965), aunque en Balcells (1964) ya se recoge esa información citando Balcells (en prensa). Por otra parte Niethammer (1964) encuentra un resto de cráneo en egagrópilas de Santander, aunque duda en atribuirlo a Myotis emarginatus o M. nattereri. En Iberia se conoció su presencia por primera vez gracias a Themido (1928) que catalogó 9 hembras procedentes de Coimbra (Portugal). El género del epíteto específico de los géneros acabados en –otis u –otus ha sido motivo de confusión a finales del siglo XX. Según Woodman 1993 Myotis es femenino y por tanto en nuestro caso debería ser emarginata para concordar con el género. Pritchard 1994 responde al artículo de Woodman corrigiendo sus afirmaciones y posteriormente Gardner 2005 argumenta que en este caso es masculino, procedente del griego: Myotis = My (ratón) + ot (oreja) + is (terminación de la segunda declinación en latín) = masculino. Además si se sigue la norma de nomenclatura zoológica de dar por bueno el nombre más antiguo se debiera mantener emarginatus. Esta situación ha supuesto un paréntesis en la parte de la producción bibliográfica entre mediados de los 90 y mediados de los 2000 en que se ha utilizado emarginata. Algo menos del 4% de los resultados de búsqueda en internet utilizando ambos términos (www.google.com, febrero 2014) corresponden a emarginata.
Origen y evolución Tradicionalmente se ha incluido en el subgénero Myotis, que incluye aquellas especies relativamente grandes con orejas largas, alas anchas, dentición menos plesiomórfica (menos primitiva), y que son típicamente cazadores sobre superficies, atrapando a sus presas sobre el suelo o la vegetación (Findley, 1972). Sin embargo las herramientas actuales de sistemática molecular han permito constatar la inconsistencia filogenética de la división del género Myotis según sus tipos ecomorfológicos (Ruedi y Mayer, 2001), siendo en principio M. dasycneme (del subgénero Leuconoe) la especie más próxima entre las europeas (Ruedi y Mayer, 2001). Sin embargo este no es un capítulo cerrado porque con sus alrededor de 100 especies y su distribución mundial, la filogenia de éste género es difícil de trabajar. Se trata del género de mamíferos terrestres con una mayor distribución natural, después de Homo (Nowak, 1994). Según las especies con las que se haya comparado en cada trabajo aparece en una posición u otra (tabla 1)
Tabla 1. Especie más próxima a Myotis emarginatus, según distintos trabajos.
Identificación Género reconocible por presentar un trago fino, recto y alargado. Se puede confundir con otros Myotis de tamaño pequeño: M. escalerai, M. bechsteinii, M. daubentoni y M. capaccinii, aunque para el ojo experimentado no será necesario capturarlo para su correcta identificación gracias a su pelaje rubio rojizo. Además M. escalerai -y el resto de especies del grupo nattereri- se diferencia por tener una franja conspicua de pelos fuertes en el borde del uropatagio a la altura del espolón; M. bechsteinii tiene unas orejas muy grandes que recuerdan un Plecotus; M. daubentoni es más oscuro y pequeño y, al igual que M. capaccini presenta unos pies muy grandes.
Descripción Murciélago de tamaño mediano-pequeño (Figura 1), con orejas de longitud media que extendidas llegan justo hasta el hocico. Presentan en la oreja una emarginación más marcada que en otras especies, aunque no es un carácter conspicuo en cualquier caso. Trago puntiagudo que apenas alcanza dicha emarginación. Pelaje de aspecto lanoso rubio rojizo dorsalmente y amarillento ventralmente. El patagio alar se inserta en la base del dedo más externo del pie. El espolón ocupa la mitad del borde posterior del uropatagio, que generalmente presenta algunos pelos pequeños. La tibia está parcialmente cubierta de pelos rojizos en su parte dorsal. (Descripción modificada de Ibáñez, 1998). Interior de la oreja con numerosas protuberancias granulares (Flaquer y Puig, 2012).
Figura 1. Myotis emarginatus. (C) J. Nogueras
Jóvenes más oscuros sin tonos rojizos. (Schober y Grimmberger, 1993). Hembras más grandes que los machos (antebrazo: 39,9-42,6 vs 37,8-39,7) (datos de los ejemplares de la colección de la EBD, en Ibáñez, 1998). Masas corporales registradas en España entre 5,5 g de un macho de Andalucía y 11,5 g de una hembra de Navarra (Ver Tablas 2 y 3). Fórmula dentaria: 2.1.3.3/3.1.3.3 (Schober y Grimmberger, 1993). Cariotipo: 2n = 44, el carácterístico de éste género (Pérez-Suarez et al., 1991).
Tabla 2. Longitud del antebrazo y peso de ejemplares adultos en diversas localidades ibero-baleares y una localidad italiana como comparación. Nótese que no se ha diferenciado entre hembras grávidas y no.
Tabla 3. Biometría general (n= 13 hembras adultas). Datos de ejemplares de Soria y Santander. (Tupinier, 1982).
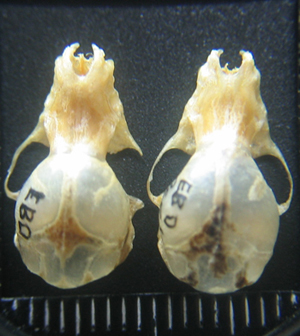
Esqueleto craneal Cráneo pequeño (ver datos españoles en Tabla 4), poco robusto, poco diferenciado de Myotis escalerai pero más alargado, de tamaño parecido a Myotis capaccinii. Rostro más alargado y estrecho que M. escalerai (figuras 1 y 2). Punta del rostro algo elevada (figura 3).
Figura 2. Vista cenital de los cráneos de Myotis emarginatus (Izquierda) y M. escalerai. Ejemplares ibéricos de la Colección de Mamíferos de la Estación Biológica de Doñana (CSIC). © J. Quetglas/M. Napal.
El canal del rostro de M. escalerai es ancho mientras que el de M. emarginatus es estrecho y muy definido (Figura 3). Estrechamiento interorbital muy marcado (Figura 2).
Tabla 4. Biometría craneal y mandibular de ejemplares ibéricos.
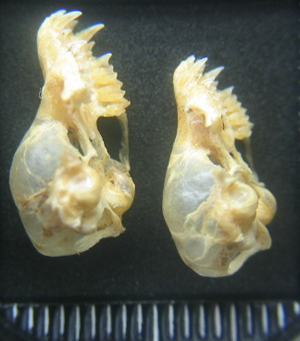
Figura 3. Vista lateral del cráneo de Myotis escalerai (izquierda) y Myotis emarginatus. Ejemplares ibéricos de la Colección de Mamíferos de la Estación Biológica de Doñana (CSIC). © J. Quetglas/M. Napal.
El arco cigomático no sobresale mucho. Inserción perpendicular a la caja craneal (figura 3). Visto lateralmente es ancho, diferenciándose del de Myotis nattereri s.l. que es más estrecho. (Kowalski y Ruprecht, 1981) Contraste marcado entre la altura de la caja craneana y el rostro en comparación con otras especies en la que la transición es gradual (Figura 4). Incisivos poderosos. (Figura 4). Bulas timpánicas pequeñas.
Figura 4. Myotis emarginatus (izquierda) y M. daubentoni. Ejemplares ibéricos de la colección de mamíferos de la Estación Biológica de Doñana (CSIC). © Juan Quetglas/María Napal.
Mandíbula Proceso coronoide triangular romo. Borde anterior casi vertical y recto. Parte posterior (superior) ligeramente convexo y un poco curvado hacia abajo (hacia el cóndilo). El proceso angular está curvado hacia fuera y se extiende sólo un poco sobre el cóndilo. Dentición (nomenclatura de los premolares: [falta P1], P2, P3 y P4): Tamaño parecido a Myotis escalerai y M. capaccinii con la formación dentaria típica del género. Maxilar superior: Todos los géneros ibéricos presentan menos de tres premolares maxilares, excepto Myotis que tiene tres y que además es el que mayor número de piezas presenta (38).P3 mucho más pequeño que P2. El pequeño premolar P3 está situado en la cara palatina de la serie dentaria. Su diámetro es mayor que en el espacio libre entre los cíngulos de P3 y P4, aunque es una morfología variable. El cíngulo de P4 presenta una cúspide en su cara mesial (la cara proximal). P4 = 3/4 de la altura de C1. Maxilar inferior: El premolar P4, muy desarrollado, presenta una sección rectangular.
Báculo Tiene la forma más simple de las especies europeas con una incisión poco visible en su basis próxima. El extremo distal es romo. No tiene cresta mediana en su superficie dorsal pero baja hacia los bordes y tiene forma como el tejado de una casa.
Ultrasonidos Utiliza pulsos de frecuencia modulada y de características variables dependiendo del entorno y la actividad. (Schumm et al., 1991). Dada esta gran plasticidad resulta muy difícil en la práctica discriminar entre las distintas especies de Myotis pequeños sin la ayuda de más información como la forma de vuelo, el comportamiento, el hábitat y el color del vientre. (Ahlén, 1990).
Variación geográfica Todas las poblaciones de Europa pertenecen a la subespecie nominal, de talla más pequeña (Ibáñez, 1998; Topál, 2001). Varios autores en sus respectivas recopilaciones (Horáček et al. 2000, Topál 2001, Simmons, 2005) han incluido otras subespcies basadas en diferencias de color y tamaño: M. e. emarginatus (Geoffroy, 1806), M. e. desertorum (Dobson, 1875 Irán), M. e. turcomanicus (Bobrinskoy, 1925, Turkmenistán), y M. e. kuzyakini (Rossolimo y Pavlinov, 1979, Uzbequistán) aunque Benda et al. (2006) consideran que el color puede no reflejar la taxonomía de la especie. La posibilidad de que existiesen especies gemelas crípticas en iberia ha sido explorada mediante análisis genético mitocondrial y nuclear con resultados negativos (Ibáñez et al., 2006)
Registro fósil En el registro fósil aparece esta especie en la península Ibérica en el Pleistoceno medio, en dos yacimientos de la provincia de Málaga (Sevilla, 1987) y M. sp. cf. emarginatus en el Pleistoceno superior y Holoceno de Ibiza (Alcover, 2003). ReferenciasAlcalde, J. T. (1995). Distribución y fenología de los quirópteros de Navarra. Tesis doctoral. Universidad de Navarra. 430 pp. Alcover, J. A. (2003). Les rates pinyades (Mammalia: Chiroptera) fossils del jaciment paleontológic del Pouás (St. Antoni de Portmany, Eivissa). Endins, 25: 141-154. Alhén. I. (1990). Identification of bats in flight. Swed. Soc. Cons. Nat. Stockholm, 50p. Balcells, E. (1964). Sobre nuevas citas de mamíferos de la Península Ibérica. Bol. R. Soc. Española Hist. Nat. (Biol.),62: 367-368. Balcells, E. (1965). Nuevos datos sobre murciélagos raros en cuevas españolas. Miscelánea Zoológica, 2: 149-160. Benda, P., Andreas, M., Kock, D., Lučan, R. K., Munclinger, P., Nová, P., Obuch, J., Ochman, K., Reiter, A., Uhrin, M., Weinfurtová, D. (2006). Bats (Mammalia: Chiroptera) of the Eastern Mediterranean. Part 4. Bat fauna of Syria: distribution, systematics, ecology. Acta Soc. Zool. Bohem. 70: 1–329. de Paz, O., Lope, M. J. (1985). Note on the distribution of the Geoffroy’s bat Myotis emarginatus (Geoffroy, 1806) in the Iberian Peninsula. Doñana, Acta Vertebrata, 12: 333-334. Findley, J. S. (1972). Phenetic relationships among bats of the genus Myotis. Syst. Zool.,21: 31-52. Flaquer, C., Puig, X. (2012). Els ratpenats de Catalunya. Editorial Brau. 141 pp. Gardner, A.L. (2005). The gender of genus-group names ending in either –otis or –otus. The Bulletin of Zoological Nomenclature, BZN 62(3). Descargado el 15-11-2007 Horáček, I., V. Hanák and J. Gaisler. 2000. Bats of the Palearctic region: a taxonomic and biogeographic review. In: (B.W. Wołoszyn, ed.) Proceedings of the VIIIth European Bat Research Symposium. Vol. I. Approaches to biogeography and ecology of bats. Chiropterological Information Center, Institute of Systematics and Evolution of Animals PAS, Kraków, Poland. pp. 11–157. Gisbert, J., Melendro, J. (1978). Datos sobre el Myotis emarginatus en la Península Ibérica. Doñana, Acta Vertebrata, 5: 113-114 Ibáñez, C. (1998). 3. Los quirópteros. Pp. 114-218. En: Blanco, J. C. (Ed.). Mamíferos de España, I. Insectívoros, Quirópteros, Primates y Carnívoros de la península Ibérica, Baleares y Canarias. Planeta, Barcelona. Ibáñez, C., García-Mudarra, J. L., Ruedi, M., Stadelman, B., Juste, J. (2006). The Iberian contribution to cryptic diversity in European bats. Acta Chiropterologica, 8 (2): 277-297. Kowalski, K., Ruprecht, A. L. (1981). Order Bats- Chiroptera. Pp. 101-154. En: Pucek, Z. (Ed.). Key to vertebrates of Poland. Mammals. Warszawa. Niethammer, J. (1964). Ein beitrag zur kenntnis der kleinsäuger nordspaniens. Z. Saugetierk., 29: 193-219. Nowak, R. M. (1994). Walker’s Bats of the World. 5th Edition. Johns Hopkins University Press, Baltimore & London, 287 pp Pérez-Suárez, G., Reina, J. M., Navlet, J., de Paz, O., López-Caballero, E. J. (1991). Citogenética de quirópteros en la península ibérica. Pp. 305-325. En: Benzal, J., de Paz, O. (Eds.). Los murciélagos de España y Portugal. ICONA – MAPA, Madrid. Pritchard, P.C.H. (1994). Comment on Gender and Declension of Generic Names. Jounal of Mammaloy 75(2): 549-550 Quetglas, J. (1997). Los murciélagos de Menorca. Informe inédito. Institut Menorquí d’Estudis, Mahón. 44 pp. Ruedi, M., Mayer, F. (2001). Molecular sistematics of bats of the genus Myotis (Vespertilionidae) suggests deterministic ecomorphological convergences. Molecular Phylogenetics and Evolution, 21 (3): 436-448. Schober, W., Grimmberger, E. (1993). Bats of Britain and Europe. Hamlyn Publising Group, London. 224 pp. Sevilla, P. (1987). Quaternary Bats of Spain: Paleoecologic and biogeographic interest. Pp. 349-355. En: Hanak, V., Horacek, I., Gaisler, J. (Eds.). European Bat Research 1987. Charles University Press, Prague. Simmons, N. B. (2005). Order chiroptera. pp. 312–529. En: Wilson, D.E., Reeder, D. M. (Eds.). Mammal species of the World: a taxonomic and geographic reference. Third edition. Vol. 1. The Johns Hopkins University Press, Baltimore, MD. Schumm, A., Krull, D., Neuweiler, G. (1991). Echolocation in the notch-eared bat, Myotis emarginatus. Behav. Ecol. Sociobiol., 28: 255-261. Stadelmann, B., Lin, L.-K., Kunz, T. H., Ruedi, M. (2007). Molecular phylogeny of New World Myotis (Chiroptera, Vespertilionidae) inferred from mitochondrial and nuclear DNA genes. Molecular Phylogenetics and Evolution, 43: 32–48 Themido, A. A. (1928). Catalogue des chiroptères existants dans les collections du Musée Zoologique de Coimbra. Mem. Est. Mus. Zool. Coimbra, 16: 5-14. Topál, G. (2001). Myotis emarginatus (Geoffroy, 1806) – Wimperfledermaus. Pp. 369-404. En: Krapp, F. (Ed.). Handbuch der Säugetiere Europas. Band 4: Fledertiere. Teil I: Chiroptera I. Rhinolophidae, Vespertilionidae 1. Aula Verlag, Wieberlsheim. Tsytsulina, K., Dick, M. H., Maeda, K., Masuda, R. (2012). Systematics and phylogeography of the steppe whiskered bat Myotis aurascens Kuzyakin, 1935 (Chiroptera, Vespertilionidae). Russian Journal of Theriology, 11: 1–20. Tupinier, Y. (1982). Quirópteros de España. Sistemática, biogeografía. Tesis doctoral presentada en 1975 en la Universidad Claude Bernard (Lyon I). Cuadernos de Espeleología. 9-10: 215-290. Tupinier, Y. (2001). Historique de la description des espèces européenes de Chiroptères. Le Rhinolophe, 15:1-140 Woodman, N. (1993). The correct gender of mammalian generic names ending in –otis. J. Mamm., 74 (3): 544-546.
Juan Quetglas Fecha de publicación: 26-03-2015 Quetglas, J. (2015). Murciélago Ratonero Pardo – Myotis emarginatus. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Barja, I. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||