|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: Iberian Mole, reproduction, demography.
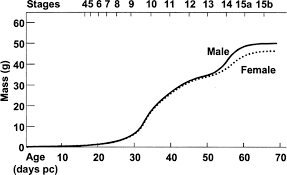
Biología de la reproducción La temporada reproductora del topo ibérico comprende los meses entre septiembre y mayo en las sierras Béticas, mientras que durante los meses de verano tiene lugar el periodo de reposo (Prochel et al., 2008). En la Cordillera Cantábrica se han encontrado hembras con embriones entre marzo y mayo, lo que sugiere reproducción primaveral (Niethammer, 1990). La gestación dura 28 días (Prochel et al., 2008). El tamaño medio de camada es de cuatro crías, aunque puede oscilar entre dos y seis (Blanco, 1998; Román, 2007), siendo estas altriciales (Román, 2007). La emancipación de las crías tiene lugar aproximadamente un mes después de su nacimiento (Barrionuevo et al., 2004a). Además, como característica exclusiva, los topos son los únicos mamíferos hermafroditas fértiles (Barrionuevo et al., 2004b). Los machos de topo ibérico presentan un periodo de actividad sexual que va desde septiembre hasta mayo, y se mantienen inactivos sexualmente el resto del año. En el periodo de septiembre a febrero se alcanzan los valores máximos de actividad sexual, medidos a través del peso testicular, del diámetro de los túbulos seminíferos y del contenido de esperma en el epidídimo. Desde mayo a agosto los machos se vuelven estériles, descendiendo el peso testicular drásticamente, alcanzando los valores mínimos durante los meses de junio y julio. Seguidamente, durante agosto y septiembre, tiene lugar un periodo de rápido desarrollo, coincidiendo con el inicio de la actividad sexual, completándose así el ciclo (Jiménez et al., 1990) (Jiménez et al., 1990). El ciclo de actividad de las hembras es paralelo. Presentan un periodo de reposo desde mayo a agosto, teniendo lugar el periodo reproductor durante el resto del año. Durante la época de cría el útero aumenta de tamaño y recibe mayor aporte sanguíneo (Jiménez et al., 1990). Pueden encontrarse hembras gestantes desde octubre hasta abril y en lactancia desde noviembre a mayo (Jiménez et al., 1990). Se trata de un periodo de actividad sexual muy largo en comparación con otros estudios llevados a cabo con Talpa europaea, los cuales evidencian una época de cría que oscila entre unos tres meses en el norte y centro de Europa (Grulich, 1967) y cinco o seis meses en la península Ibérica (López-Fuster et al., 1988). Aunque estos estudios implican a dos especies diferentes, los resultados sugieren la existencia de un gradiente latitudinal que afecta a la longitud del periodo de cría, acortándose en las poblaciones del norte y ampliándose gradualmente a medida que nos acercamos al ecuador. Este hecho podría deberse a las diferencias en las condiciones climáticas en el norte y sur de Europa (López-Fuster et al., 1988; Jiménez et al., 1990; Blanco, 1998). Hermafroditismo Los topos poseen una característca única dentro de los mamíferos y es que las hembras de las especies Talpa occidentalis, T. europaea,T. romana y T. stankovici presentan ovotestis bilaterales en lugar de ovarios (Jiménez et al., 1993; Sánchez et al., 1996). La parte ovárica contiene oocitos maduros durante el periodo reproductor y es por tanto funcional. La parte testicular presenta un tamaño variable y no contiene células germinales. Esta ultima está compuesta por un tejido testicular anormal que contiene numerosas estructuras similares a los túbulos seminíferos pero demasiado pequeñas o esféricas, en una densa matriz de células de Leydig, las cuales producen testosterona. Las células de Sertoli presentan características fetales y no están acompañadas de células germinales. En los ovotestis también se encuentra un rudimentario epidídimo (Jiménez, et al., 1993; Jiménez, et al., 1996). El hermafrofitismo dentro del género Talpa se mantiene prácticamente sin alteraciones a través del periodo evolutivo de divergencia entre las diferentes especies del género. De hecho, todas las características anatómicas, histológicas y moleculares descritas en T. occidentalis (Jiménez et al., 1993) se encuentran de igual modo en las otras tres especies del género estudiadas (T. europaea,T. romana y T. stankovici). Estas características sugieren que una mutación o mutaciones genéticas fueron responsables del desarrollo del tejido testicular en las gónadas de las hembras de esta especie. Las hembras de topo son hermafroditas y presentan un genotipo XX, por lo que carecen del gen SRY que está ligado al cromosoma Y y es el responsable de la determinación del sexo masculino. Esta mutación permite la activación de otros genes implicados en la determinación del sexo sin la intervención del gen SRY (Sánchez et al., 1996). Debido a que esta característica parece encontrarse de forma generalizada entre las especies del género Talpa probablemente se trate de una mutación muy antigua, que fue fijada en las poblaciones de topos que dieron lugar a las especies actuales (Sánchez et al., 1996). Esfuerzo reproductivo En las hembras de topo ibérico, el coeficiente de esfuerzo reproductivo materno, definido como la masa de la descendencia en el destete en relación con la masa de la madre, es inversamente proporcional a la masa materna (Barrionuevo et al., 2004a). Comparados con otros insectívoros soricomorfos de menor masa corporal (musarañas), en los topos dicho coeficiente (2.69) es significativamente más alto que el esperado (1.91), lo que conlleva que su esfuerzo reproductivo sea un 36,4% mayor (Barrionuevo et al., 2004a). Este dato implica que las hembras de topo presentan una increíble capacidad de alimentación de las crías en comparación con otros insectívoros. Como consecuencia, las crías de topo están casi completamente desarrolladas en el momento del destete, hecho que podría contribuir a disminuir la tasa de mortalidad durante este periodo crítico (Barrionuevo et al., 2004a). Niveles de testosterona Se han encontrado grandes variaciones en los niveles de testosterona en sangre tanto en machos como en hembras, dependiendo de la edad y de la fecha de captura. Esta variación es tan grande incluso dentro de los grupos que poco puede concluirse acerca de ello. En cualquier caso, los niveles de testosterona en los machos se corresponden con el ciclo actividad sexual, mientras que en las hembras ocurre lo contrario, durante la época no reproductora este nivel es similar al de los machos, mientras que durante los meses de la temporada de cría es mucho menor (Jiménez et al., 1993). Sorprendentemente los niveles de testosterona se correlacionan mejor con el peso del ovotestis que con el peso testicular de los machos (Jiménez et al., 1993). Esta correlación sugiere que la testosterona encontrada en las hembras es producida por las células de Leydig presentes en el tejido testicular del ovotestis (Jiménez et al., 1993). Desarrollo y crecimiento Se han establecido 15 estadios que comprenden todo el desarrollo de T. occidentalis (Figura 3) (Barrionuevo et al., 2004a). El desarrollo prenatal dura hasta el estadio 8 (s8), tras el nacimiento, cada 5 días se alcanza un nuevo estadio, correspondiendo s15 a juveniles completamente desarrollados (Barrionuevo et al., 2004a). Tanto los machos como las hembras de topo ibérico presentan una curva trifásica de crecimiento (Figura 3) (Barrionuevo et al., 2004a). Estas tres fases están marcadas por eventos significativos en el desarrollo (Barrionuevo et al., 2004a). La primera fase de crecimiento coincide con el periodo prenatal y, por lo tanto, termina con el nacimiento, que tiene lugar a los 28 días del coito (Prochel et al., 2008). En la primera parte de esta fase (s1-s6), aunque constituye un periodo importante en el cual los órganos y tejidos se diferencian, no tiene lugar un intenso crecimiento. Sin embargo, durante el desarrollo prenatal tardío (s7-s9), el crecimiento predomina sobre los procesos de diferenciación y organogénesis. La segunda fase comienza con la mayor tasa de crecimiento (s10), probablemente debido a un alto contenido en nutrientes en la leche materna, y termina con un periodo de baja tasa de crecimiento que coincide con la formación del pelo (s12). Tras este, un nuevo aumento en la tasa de crecimiento marca el comienzo de la tercera fase, la cual termina con las crías alcanzando el tamaño juvenil (s15). El estadio 15 se subdivide en dos para diferenciar a las crías que permanecen en el nido (s15a) de las que han sido destetadas (s15b). Los topos juveniles sufren una pérdida de peso corporal tras el destete debida a la repentina necesidad de buscar su propio alimento y territorio. Este periodo es crítico y presenta una elevada tasa de mortalidad. Los juveniles que sobreviven establecen su propio territorio y el crecimiento puede reanudarse recuperando el peso perdido (Barrionuevo et al., 2004a). Los juveniles (s15b) se mantienen sin importantes cambios fisiológicos hasta que alcanzan la pubertad al comienzo de la siguiente temporada reproductora, por lo que este último estadio puede prolongarse hasta 10 meses en el caso de T. occidentalis (Barrionuevo et al., 2004a).
Figura 1. Incremento de la masa corporal del topo ibérico (Talpa occidentalis) durante los 15 estadios de su desarrollo, mostrando una curva trifásica de crecimiento. La edad es reflejada en días postcoito (Barrionuevo et al., 2004a).
Estructura y dinámica de poblaciones Los machos de topo ibérico no alcanzan la madurez sexual hasta aproximadamente un año de edad, antes de este punto, sus gónadas muestran características típicamente juveniles (Jiménez et al., 1990).
Referencias Barrionuevo, F. J., Zurita, F., Burgos, M., Jiménez, R. (2004a). Developmental Stages and Growth Rate of the Mole Talpa occidentals (Insectivora, Mammalia). Journal of Mammalogy, 85 (1): 120-125. Barrionuevo, F. J., Zurita, F., Burgos, M., Jiménez, R. (2004b). Testis-like development of gonads in female moles. New insights on mammalian gonad organogenesis. Developmental biology, 268 (1): 39-52. Blanco, J. C. (1998). Mamíferos de España. I. Insectívoros, Quirópteros, Primates y Carnívoros de la península Ibérica, Baleares y Canarias. Geo Planeta Guías de campo, Barcelona. 457 pp. García-López de Hierro, L. G. L., Moleón, M., Lupiáñez, D. G., Virgós, E., Jiménez, R. (2013). Positive and negative unintended human-induced effects on Iberian mole abundance at the edge of its distribution area. Mammalian Biology-Zeitschrift für Säugetierkunde, 78 (4): 276-282. Grulich, I. (1967). Die Variabilität der taxonomischen Merkmale des Maulwurfs (Talpa europaea L., Insectivora) im zusammenhang mit Alter und Geschlecht. Zool. Listy, 16: 125-144. Jiménez, R., Alarcón, F. J., Sánchez, A., Burgos, M., de la Guardia, R. D. (1996). Ovotestis variability in young and adult females of the mole Talpa occidentalis (Insectivora, Mammalia). Journal of Experimental Zoology, 274 (2): 130-137. Jiménez, R., Burgos, M., Sánchez, A., de La Guardia, R. D. (1990). The reproductive cycle of Talpa occidentalis in the southeastern Iberian Peninsula. Acta Theriologica, 35 (1-2):165-169. Jiménez, R., Burgos, M., Sánchez, A., Sinclair, A. H., Alarcón, F. J., Marín, J. J., Ortega, E., de la Guardia, R. D. (1993). Fertile females of the mole Talpa occidentalis are phenotypic intersexes with ovotestes. Development, 118 (4): 1303-1311. López-Fuster, M. J., Gosálbez, J., Lluch, S. (1988). Characteristics of the Reproductive Cycle of the Mole, Talpa europaea, in the Northeast of the Iberian Peninsula. Acta Theriologica, 33 (11): 131-137. Prochel, J., Goswami, A., Carmona, F. D., Jimenéz, R. (2008). Ossification sequence in the mole Talpa occidentalis (Eulipotyphla, Talpidae) and comparison with other mammals. Mammalian Biology-Zeitschrift für Säugetierkunde, 73 (5): 399-403. Román, J. (2007). Talpa occidentalis Cabrera 1907. Pp. 89-91. En: Palomo, L. J., Gisbert, J., Blanco, J. C. (Eds.). Atlas y Libro Rojo de los Mamíferos Terrestres de España. Dirección General para la Biodiversidad-SECEM-SECEMU, Madrid, 588 pp. Sánchez, A., Bullejos, M., Burgos, M., Hera, C., Stamatopoulos, C., de la Guardia, R. D., Jiménez, R. (1996). Females of four mole species of genus Talpa (Insectivora, Mammalia) are true hermaphrodites with ovotestes. Molecular reproduction and development, 44 (3): 289-294.
Mª Carmen Hernández González Fecha de publicación: 11-07-2016 Hernández, M. C. (2016). Topo ibérico – Talpa occidentalis. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Barja, I. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|
|
|
|
|
|