|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: Rufous Bush Robin, reproduction, nest quality, egg size, breeding success, senescence.
Biología de la reproducción Emparejamiento Los machos cantan con una tasa máxima y más cerca del nido durante el periodo de establecimiento del territorio y formación de la pareja (Álvarez, 1996b). Cantan situados en elementos elevados del ambiente (López, 1983), entre 0,5 y 8 m de altura (Valverde, 1957). A veces realizan vuelos de canto (Beven, 1970). Posteriormente, la tasa de canto disminuye en el periodo fértil de la pareja y en las fases reproductivas posteriores (Álvarez, 1996b). La tasa de canto también disminuye con el avance de la fecha de puesta (Álvarez, 1996b). Por tanto, la función principal del canto es la adquisición de la pareja y el establecimiento en un territorio (Álvarez, 1996b), pero también es usado para buscar cópulas extrapareja (Álvarez, 1997). Existe una correlación entre el tamaño de las marcas blancas terminales de la cola dentro de la pareja, y también respecto al tamaño corporal (marginalmente no significativa; Álvarez, 2004a). Esto sugiere que las manchas blancas de la cola pueden intervenir en los procesos de emparejamiento, ya que la cola es exhibida en los cortejos (Álvarez, 2000b). Es una especie muy territorial, con territorios de 2 a 8 ha de tamaño (López, 1983), con una media de 2,36 ha (López y Gil-Delgado, 1988). En el Sáhara, los machos se separan al menos 200 m entre ellos (Valderde, 1957). Son monógamos. Se ha descrito sólo un posible caso de poligamia, al haberse encontrado dos nidos separados 125 m entre sí con un sólo macho visto en las inmediaciones (López y Gil-Delgado, 1988). Nidificación El alzacola nidifica en diferentes cultivos, sobre todo en vides y olivos, pero también utiliza otras especies leñosas o incluso nidifica directamente sobre el suelo en huecos en los árboles o en construcciones humanas (Cano, 1960; López, 1983; López y Gil-Delgado, 1988; Palomino et al., 1998c). También ocupa pinos de porte arbustivo, que se encuentran próximas al litoral (López, 1989). En Sevilla, nidifica en vides a una altura media de 0,5 m (Palomino et al. 1998c). En Alicante, el nido se sitúa a una altura media de entre 1 y 2 m. Se sitúan a mayor altura cuanto más alto es el árbol (López, 1989). López (1983) reporta una media de altura de 2,1 m (rango 1,7-2,5 m). En olivos, el alzacola prefiere una orientación noroeste (NO), para estar en umbría y evitar un sobrecalentamiento en verano (López, 1989). Ambos sexos intervienen en la construcción del nido, que es relativamente grande para el tamaño del ave, y poco compacto, por lo que utiliza ramas gruesas que lo sujeten (López, 1989). El nido está formado por una taza de material vegetal (ramas y hierba), con la cavidad central acolchada con fibras, algodón, plumas, pelo y raíces (Harrison, 1991). El diámetro del nido y la profundidad varía según la zona y estudios realizados. López (1983) describe que el diámetro exterior es de 13 a 20 cm, el interior de 7-9 cm, y la profundidad del nido de 4 a 6,5 cm. En Alicante, los nidos tienen un diámetro exterior de 17-19 cm, diámetro del cuenco de 7 cm, altura de 11 cm, y profundidad del cuenco de 5 cm, con un peso promedio de 75 gr (López, 1989). En Sevilla, tiene 13,4 cm de diámetro (rango= 9-17,5), pesa una media de 105 gr (rango= 34-226 g), y tiene una profundidad de la copa de 4,4 cm (rango 2,5 a 5,8) (Palomino et al., 1998c). En Argelia, el tamaño del nido es mayor cuanto más alto está en la vegetación y el éxito reproductor aumenta con la altura del nido (Tabib et al., 2016). Estos autores consideran que, dado que la depredación es alta en el alzacola (más información en el apartado “Interacciones”) los nidos grandes pueden atraer a depredadores (Palomino et al., 1998c), por lo que sólo construyen nidos grandes a mayor altura (más a salvo de los depredadores). Palomino et al. (1998c) testaron diversas hipótesis para entender la evolución del tamaño del nido en el alzacola. Encontraron que la tasa de parasitismo por el cuco (Cuculus canorus) y la tasa de depredación tienden a ser menor cuanto mayor es el nido. El tamaño de puesta es mayor cuanto más material es usado en la construcción del nido, que también se correlaciona positivamente con la cantidad de alimento llevado a los pollos por el macho, y el número de volantones producidos tiende a aumentar con el peso del nido, por lo que concluyen que el tamaño del nido es usado como una señal sexual post-emparejamiento. También encontraron que la masa del nido disminuyó con la fecha de puesta. Puesta e incubación de los huevos La reproducción es tardía, usualmente empezando en el mes de mayo o incluso a primeros de junio (Harrison, 1991). En Alicante la media de la fecha de puesta del primer huevo es el 6 de junio (López y Gil-Delgado, 1988). Suele tener sólo una puesta, aunque ponen dos en algunas ocasiones (dos tercios de las parejas en el levante, López y Gil-Delgado, 1988), así como puestas de reposición. El tamaño de puesta oscila entre 2 y 5 huevos. Aunque Harrison (1991) habla de tamaños normalmente de 4-5 huevos, en España la moda suele ser 3-4 huevos, siendo menos común puestas de 2 o 5 (López y Gil-Delgado, 1988). La media es de 3,7 huevos en Alicante (López, 1983; López y Gil-Delgado, 1988). Similares tamaños de puesta se encuentran en el norte de África (Boukhriss y Selmi, 2010; Adamou et al., 2017). En Sevilla, el tamaño modal de puesta fue de 3 huevos, de los que nacieron 2-3 pollos (Palomino et al., 1999) y el tamaño de puesta fue mayor en las primeras puestas (3,5) que en las segundas o en las de reposición (3,0). Consecuentemente, también fue mayor el tamaño de nidada (3,06) frente a 2,64 (segundas) y 2,48 (reposición). Los tamaños de puesta no difieren entre primeras y segundas puestas en Alicante (López y Gil-Delgado, 1988). El tamaño de puesta también descendió con la fecha de puesta en Sevilla (Palomino et al., 1999) y en Argelia (Adamou et al., 2017). Las hembras que empezaron a reproducirse antes intentaron más puestas, pero no lograron tener mayor éxito reproductor que las más tardías. Los huevos son blancos, a veces tenuamente azulados o verdosos, muy densamente manchados con motas pardo purpúreo de diferente tamaño, normalmente más concentradas en el polo ancho (Harrison, 1991). Los huevos miden 22,5 × 16,3 mm (19,5-26 × 14,3-18 mm) y pesan 3,2 g (Cramp et al., 1998). En Alicante miden 21,3 × 16 mm (López y Gil-Delgado, 1988). Álvarez (1994a) reporta que el 16% de los huevos tienen las motas muy esparcidas y un 19% muestra las motas concentradas en una mitad más ancha del huevo. La incubación es realizada exclusivamente por la hembra, y empieza con la puesta del último huevo (López, 1983). El macho la alimenta durante ese periodo. La incubación dura una media de 13 días (rango 12-14; López y Gil-Delgado, 1988) y el último huevo eclosiona un día después de los anteriores (López y Gil-Delgado, 1988). Cuidado de los pollos El pollo es nidícola y nace desnudo, con piel oscura, boca naranja-amarillo y comisuras blancas (Harrison, 1991). Los polluelos permanecen en el nido entre 8 y 13 días, con una media de 10,6 días (López y Gil-Delgado, 1988; Álvarez, 1994c). Alcanzan la asíntota de crecimiento a los 8 días (López y Gil-Delgado, 1988), siguiendo la fórmula: Peso = 21,56/(1+e-0,562(t-3,44)). Los pollos de alzacola abren los ojos a los 6 días (López, 1983). A los 7 días ya dan señales de miedo, de modo que al ser tocados se tumban todo lo que pueden en el fondo del nido. Moreno-Rueda et al. (2009) estudió en detalle la alimentación de los pollos. Las hembras ceban más que los machos (9 cebas por hora frente a 5,6 los machos; ver también Martín-Gálvez et al., 2005), pero el tamaño de presa no difiere entre sexos. La edad de la nidada no afectó a la tasa de cebas ni al tamaño de las presas. El tamaño de las presas no varió con la fecha, pero la tasa de cebas sí disminuyó con la fecha. La tasa de cebas aumentó con el número de pollos que pedían en el nido, pero este efecto ocurrió solo en las hembras, las cuales responden más a las llamadas de petición de los pollos que los machos. Los padres cebaron principalmente a los pollos que pidieron con mayor intensidad, más rápido tras la llegada de los padres, más cerca de ellos, y que alcanzaron mayor altura al pedir. Los pollos más grandes recibieron más comida, debido a que solicitaron alimento más veces cerca de los padres. Es decir, los pollos más grandes perecen tener una ventaja competitiva, gracias a la cual consiguen un mejor acceso a las posiciones preferenciales para ser alimentados, y gracias a eso consiguen más comida. También tendieron a pedir más rápido, a mayor intensidad y alcanzar más altura. Después de ser alimentados, el pollo que consume la presa, pide más lejos, más tarde, a menor altura y menor intensidad en la ceba siguiente, indicativo de que estas variables están influenciadas por el nivel de hambre. Las hembras emplearon en el nido el triple de tiempo inspeccionando a los pollos antes de alimentarlos que los machos (37 frente a 14 s.). Una vez que los pollos vuelan del nido, el esfuerzo parental se divide de manera que cada padre se encarga de unos pollos concretos (en nidadas con varios pollos). Los volantones solicitan alimento de manera vistosa con llamadas y agitando las alas (López, 1983). Típicamente, se esconden en el matorral para ocultarse de posibles depredadores (López, 1983). Éxito reproductivo En una población de alzacolas en Alicante, la supervivencia de los polluelos fue similar en la primera y segunda puesta, debido a la diferencia en el éxito de eclosión, ya que las segundas puestas produjeron un mayor número de polluelos (López y Gil-Delgado, 1988). El 61% de los huevos eclosionaron, el 63% de los pollos volaron y en total un 39% de los huevos produjeron un volantón. Casi la mitad de las primeras puestas fueron destruidas por depredadores, y sólo un tercio de estas fueron exitosas (López y Gil-Delgado, 1988). El éxito reproductor tiende a ser mayor en las segundas puestas (López y Gil-Delgado, 1988). En total, López y Gil-Delgado (1988) encontraron que el 8% de los huevos que fracasaron fueron infértiles, un 2% de los pollos murieron por inanición y el 69% fueron depredados (dos tercios en estado de huevos y un tercio como pollos). Palomino et al. (1999) estudiaron la reproducción del alzacola en Sevilla, donde reportaron entre una a cuatro puestas por año, aunque sólo suelen tener una puesta exitosa, con 0 a 7 volantones producidos en total, normalmente 2 o 3 (en promedio 2,56 pollos), con una productividad de 1,26 pollos por intento reproductor. Eclosionaron el 61% de los huevos. El 59% de los huevos eclosionaron en la primera puesta, un 57% en las de reposición, y un 77% en la segunda puesta. El 61% de los pollos llega a volar, en las primeras puestas volaron un 61%, mientras que en las segundas un 50% y un 66% en puestas de reposición. Las pérdidas fueron principalmente por depredación de huevos y parasitismo de cría. El parasitismo de cría afectó sobre todo a las primeras puestas y a las puestas de reposición, mientras que la depredación de los pollos afectó a las segundas puestas. El 35% de las puestas fueron depredadas, un 25% cuando contenían huevos y un 10% con pollos. Un 17% fueron parasitados y un 5% abandonados. Además, detectaron un nido en el que los pollos murieron por inanición. En Sevilla, la masa de los pollos volantones fue entre 17,9 y 19,8 g, según el año. El número de bandas de crecimiento en la hembra se correlacionó negativamente con su éxito reproductor, mientras que la condición física (medida como residuos del peso respecto al tarso) no afectó al éxito reproductor (Palomino et al., 1999). Las hembras de primer año tuvieron puestas más tardías que las demás, además de una menor productividad y volantones con un peso menor. El divorcio no pareció afectar a la productividad. La depredación del nido no pareció afectar a la tasa de divorcio, pero los individuos que se divorciaron tendieron a disminuir su tasa de depredación en el siguiente evento reproductor, pero no aumentaron su productividad total (Palomino et al., 1999). La depredación es del 39% según Álvarez (2000b). Pueden tener de 1 a 6 sucesivos intentos y el éxito reproductor aumenta cuando es más temprana la cría de los pollos (Álvarez, 2000b). Además, el éxito reproductivo también es mayor en los alzacolas que presentan una mayor simetría en las marcas negras de la cola (Álvarez, 2000b), principalmente porque sus nidos fueron menos depredados. Esto se debe a que la cola es utilizada en un comportamiento de distracción ante los depredadores. Los alzacolas mueven la cola arriba y abajo en estos encuentros, exponiendo particularmente las marcas blancas y negras (Álvarez, 2006).
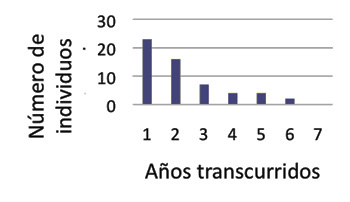
Estructura y dinámica de las poblaciones Se estudió la probabilidad de supervivencia en una población sevillana, analizando los individuos anillados y recuperados el periodo 1984-1993. La tasa interanual de supervivencia fue similar a la de otros paseriformes, de 0,57 (Domínguez y Cuadrado, 1994). Las recapturas fueron mayores en adultos (21%) que en volantones (2%). Los machos se recapturaron más frecuentemente que las hembras (35 frente al 22%). Se cree que viven como máximo 6 años (Figura 1).
Figura 1. Número de alzacolas recapturados en función de los años transcurridos. Según Domínguez y Cuadrado (1994).
Referencias Adamou, A. E., Tabib, R., Kouidri, M., Ouakid, M. L., Gl?dalski, M., Ba?bura, A., Ba?bura, J. (2017). Inter-annual variation in clutch size and laying date of Rufous Bush Chats Cercotrichas galactotes inhabiting an Algerian oasis. Journal of Arid Environments, 141: 40-44. Álvarez, F. (1994a). A gens of Cuckoo Cuculus canorus parasitizing Rufous Bush Chat Cercotrichas galactotes. Journal of Avian Biology, 25: 239-243. Álvarez, F. (1994c). Rates of weight increase of Cuckoo (Cuculus canorus) and host (Cercotrichas galactotes) chicks. Ardeola, 41: 63-65. Álvarez, F. (1996b). Variation in song rate during the breeding cycle of the Rufous Bush Chat, Cercotrichas galactotes. Ardeola, 43:49-56. Álvarez, F. (1997). The functions of song and the spatial pattern of song production in the Rufous Bush Chat (Cercotrichas galactotes). Doñana, Acta Vertebrata, 24: 67-78. Álvarez, F. (2000b). Relationship between tail color pattern and reproductive success, mate acquisition and nest predation in Rufous Bush Chats. Condor, 102: 708-712. Álvarez, F. (2004a). Black and white tail markings in rufous bush chats Cercotrichas galactotes: size, symmetry and the extent of seasonal abrasión. Ardeola, 51: 169-175. Álvarez, F. (2006). Defensa del nido por el alzacola. Pp. 89-91. En: Soler, M. et al. (Eds.). Fauna en Acción. Lynx, Barcelona Beven, G. (1970). Studies on less familiar birds. 160. Rufous Bush Chat. British Birds, 63: 294-299. Boukhriss, J., Selmi, S. (2010). Risk-taking by incubating rufous bush robins Cercotrichas galactotes: season-dependent incubation stage effect. Journal of Ethology, 28: 331-337. Cano, A. (1960). Sobre nidificación de Erythropygia galactotes en Pechina, Almería. Ardeola, 6: 320-323. Cramp, S., Simmons, K. E. L., Perrins, C. M. (1998). The Complete Birds of the Western Palearctic on CD-ROM. Oxford University Press, Oxford. Domínguez, F., Cuadrado, M. (1994). Recurrencia y tasa de supervivencia en el Alzacola Cercotrichas galactotes. Butletí del Grup Catalá d’Anellament, 11: 45-49. Harrison, C. (1991). Guía de campo de los nidos, huevos y polluelos de las aves de España y Europa. Omega, Barcelona. López, G. (1983). Datos sobre la nidificación del Alzacola (Cercotrichas galactotes). Alytes, 1: 373-392. López, G. (1989). Variación de la colocación y orientación del nido del Alzacola (Cercotrichas galactotes) en dos especies de árboles. Doñana, Acta Vertebrata, 16: 57-67. López, G., Gil-Delgado, J. A. (1988). Aspects of the breeding ecology of Rufous Bush Robins Cercotrichas galactotes in southeast Spain. Bird Study, 35: 85-89. Martín-Gálvez, D., Soler, M., Soler J. J., Martín-Vivaldi, M., Palomino, J. J. (2005). Food acquisition by common cuckoo nestlings in rufous bush robin nests and the advantage of the eviction behaviour. Animal Behaviour, 70: 1313–1321. Moreno-Rueda, G., Soler, M., Martín-Vivaldi, M., Palomino, J. J. (2009). Brood provisioning rate and food allocation rules according to nestling begging in a clutch-adjusting species, the Rufous-tailed Scrub-robin Cercotrichas galactotes. Acta Ornithologica, 44: 167–175 Palomino, J. J., Martín-Vivaldi, M., Soler, M., Soler, J. J. (1998c). Functional significance of nest size variation in the Rufous Bush Robin Cercotrichas galactotes. Ardea, 86: 177-185. Palomino, J. J., Martín-Vivaldi, M., Soler, M. (1999). Determinants of reproductive success in the rufous bush robin (Cercotrichas galactotes). Journal für Ornithologie, 140: 467-480. Tabib, R., Adamou, A. E., Ouakid, M. L., Gl?dalski, M., Ba?bura, J. (2016). Nesting height and nest-cup volume variation in Rufous Bush Chats (Cercotrichas galactotes) inhabiting Algerian oases: consequences for breeding success. Avian Biology Research, 9: 114-119. Valverde, J.A. (1957). Aves del Sahara español: estudio ecológico del desierto. Consejo Superior de Investigaciones Científicas, Madrid.
Gregorio Moreno-Rueda Inmaculada Abril-Colón Fecha de publicación: 14-12-2018 Moreno-Rueda, G., Abril-Colón, I. (2018). Alzacola rojizo – Cercotrichas galactotes. En: Enciclopedia Virtual de los Vertebrados Españoles. López, P., Martín, J., Morales, M. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org
|
|
|
|
|
|
|