|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: Leonese Rock Lizard, identification, description, measurements, variation.
Origen y evolución De acuerdo con los más recientes análisis filogenéticos (Arribas et al., 2006), I. galani pertenece al llamado “grupo ibérico” (o “grupo monticola”), que se originó hace unos 8,7 millones de años e incluye además de I. monticola, a I. martinezricai e I. galani (diferenciadas entre sí hace unos 2,5 millones de años). Además, en este “grupo ibérico” se incluye la lagartija carpetana (I. cyreni) que está bien diferenciada dentro de él (hace unos 7,5 millones de años). La filogenia sugiere que I. monticola podría ser el taxon hermano de I. galani de la cual se habría separado hace aproximadamente 2 millones de años, al inicio del Pleistoceno. El “grupo monticola” estuvo probablemente ampliamente distribuido por el oeste ibérico durante las fases más frías del Pleistoceno, quedando acantonado en las zonas que actualmente ocupa como resultado del incremento térmico durante el Holoceno y por competencia con otros lacértidos (Podarcis). Iberolacerta galani es endémica de los Montes de León, donde está aislada del resto de especies del “grupo monticola” por los rios Duero y Miño-Sil, pero especialmente de las poblaciones más cercanas de I. monticola, por el marcadamente mediterráneo valle del río Bibei. El factor clave en la separación (casi sincrónica) de estas tres especies del “grupo monticola” se da hacia el final del Mioceno (fase antíllica), momento en el cual toda esta área general se eleva y la red hidrográfica se encaja aprovechando el relieve preexistente. Más tarde, durante la fase rodánica (ya en el Plioceno) la fosa del Bierzo se hunde y las cuencas de los ríos Eria y Sil aparecen ya tal y como hoy en día las conocemos, tal y como ocurrió con otros ríos del oeste ibérico (como la cuenca del Duero, ver Arribas y Carranza, 2004), actuando como barrera y desencadenando la prácticamente simultánea especiación entre I. galani, I. martinezricai e I. monticola. En el caso de I. martinezricai, la basculación de la península Ibérica hacia el oeste abrió hacia el océano Atlántico la previamente endorreica cuenca del Duero (Arribas y Carranza, 2004) aislando a esta especie al sur de dicho río. También en I. galani, esta misma basculación hacia el oeste podría haber dado lugar a la captura de las partes altas del rio Bibei, que cambió su dirección y en vez de drenar hacia el río Tera (es decir, hacia la cuenca del Duero), comenzó a drenar hacia el oeste, y después hacia el norte, hacia la cuenca del Miño-Sil, cortando el posible corredor este-oeste de Iberolacerta en el área, y dejando a I. galani, ya confinada entre los ríos Duero y Miño-Sil, además, aislada de otras I. monticola más occidentales por el marcadamente mediterráneo valle del rio Bibei. El refugio würmiense de I. galani podría haber sido más o menos “in situ”, ya que los datos genéticos sugieren que el aislamiento con I. monticola (presente apenas a 40 km de distancia) no se ha roto a lo largo del Cuaternario. Este refugio podría haber estado situado en alguna zona más o menos cercana a su área actual, la cual, no obstante, permaneció fuertemente glaciada durante las fases más frías del Würm. Este refugio podría haber estado situado tanto hacia el norte de su área actual (en el Bierzo, pero siempre al sur del río Sil, que es una barrera) o más probablemente hacia el sur de los Montes de León (entre las montañas que ocupa la especie actualmente y el río Duero, donde existen actualmente áreas de clima mesomediterráneo, con especies marcadamente termófilas). Iberolacerta galani puede considerarse un “reinmigrante a corta distancia” (sensu Holdhaus, 1902, 1906, 1954) originado más o menos "in situ", tal y como ocurre con otras especies endémicas del área. Un mecanismo similar ha sido sugerido para la especiación a muy escasa distancia de las tres especies pirenaicas de Iberolacerta (Pyrenesaura) (Mayer y Arribas, 1996; Carranza et. al. 2004).
Diagnosis Diagnosis general Iberolacerta de talla grande, especialmente caracterizada por la siguiente combinación de caracteres (Arribas et al., 2006; Arribas, 20142): Longitud cabeza-cloaca notable (hembras hasta 84,42 mm, la mayor Iberolacerta conocida hasta la fecha es una hembra de esta especie) con patas anteriores y posteriores comparativamente más cortas. Numerosos ocelos axilares azules (reflectantes en UV, como los puntos azules de las ventrales más externas). Contacto entre las escamas supranasal y loreal relativamente frecuente (contacto pleno en cerca de un cuarto de los especímenes estudiados, y cerca de contactar en muchos otros). Además, tiene una frecuencia relativamente baja de contacto rostral-internasal en los machos (33 %) pero relativamente más alto en las hembras (58 %), mayor número de collaria, dorsalia y ventralia y menor de circumanalia en comparación con otras Iberolacerta, aunque los valores de folidosis parecen reflejar factores climáticos de las localidades en cuestión. Escamas postocular y parietal separadas. Escama supernumeraria (“azygos”) entre las prefrontales rara (en un 13 % de los ejemplares). Osteológicamente, se caracteriza por su escamoso bastante rectilíneo, solo incurvado en su parte posterior. Cariotipo con 36 macrocromosomas acrocéntricos (de un solo brazo) gradualmente decrecientes en talla. Organizadores nucleolares (NOR) en posición telomérica de un cromosoma grande (posiblemente el quinto par; Tipo-L). Cromosomas sexuales diferenciados y heteromórficos, presentando Z una banda peritelomérica que se tiñe con CromomicinaA 3, carácter único entre las Iberolacerta, y de tamaño grande, como el sexto par de autocromosomas; mientras que el cromosoma sexual W está heterocromatinizado y es tan largo como los pares autosómicos 10 o 11. Las secuencias de DNA mitocondrial parciales del citocromo b y de rRNA del 12S secuenciadas son distintas de los demás representantes de Iberolacerta conocidos hasta la fecha (Arribas et al., 2006). Diagnosis diferencial (las especies del “grupo ibérico” con diferencias significativas respecto a I. galani se dan entre paréntesis; abreviaciones, cyr: I. cyreni; mart: I. martinezricai; mont: I. monticola). Los caracteres que diferencian a I. galani de I. monticola (la especie más similar) han sido subrayados para mayor claridad (Arribas et al., 2006). Los machos de I. galani tienen menos gránulos supraciliares en el lado izquierdo (cyr), mayor collaria (cyr), mayor ventralia (cyr), menor femoralia en el lado izquierdo (mart), menos circumanalia (cyr), menor frecuencia de contacto entre la rostral y la internasal (cyr, mont), mayor frecuencia de contactos entre la supranasal y la loreal (todas las otras spp.), mayor número de ocelos azules (todas las otras spp.), mayor puntuación ventral (cyr), menor longitud de las patas anteriores (cyr, mont), menor longitud de la escama parietal (cyr), masetérica mayor (cyr, mart), timpánica mayor (cyr) y placa anal menor (cyr). Las hembras de I. galani tienen menos gránulos supraciliares en el lado izquierdo (cyr), mayor collaria (todas las otras spp), mayor dorsalia (cyr, mont), mayor ventralia (todas las otras spp.), menor circumanalia (cyr, mont), menor frecuencia de contacto rostral-internasal (cyr), mayor frecuencia de contacto entre la supranasal-loreal (todas las otras spp), mayor número de ocelos azules (todas las otras spp), mayor puntuación ventral (cyr), menor longitud de las patas anteriores (cyr), menor longitud de las patas posteriores (cyr, mont), píleo corto (cyr), masetérica mayor (mart), timpánica mayor (todas las otras spp.) y placa anal menor (cyr). La presencia de una escama supernumeraria entre las prefrontales es rara en I. galani (13 %), mientras que es muy común en las cercanas poblaciones gallegas de I. monticola (presente en más del 50% de los especúmenes analizados de la mayor parte de las poblaciones gallegas, y especialmente en los ejemplares de Cabeza de Manzaneda (Orense), la población más cercana conocida al área de I. galani, situada a menos de 40 km (Arribas et al., 2006).
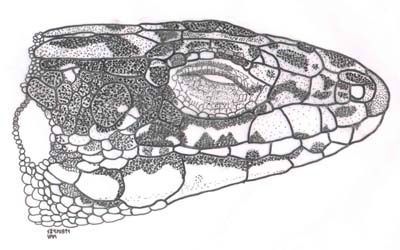
Figura 1. Aspecto lateral de la cabeza de Iberolacerta galani. © O. Arribas.
Descripción Ver apartado de diagnosis. Biometria y folidosis Machos: Basado en 24 ejemplares, se da la media ± desviación estandar (mínimo-máximo) (Arribas et al., 2006). Medidas: Longitud corporal (SVL): 60,78 ± 1,38 (45,83-69,1); Longitud de los miembros anteriores (FLL): 20,73 ± 0,58 (15,4-24,9); Longitud de los miembros posteriores (HLL): 30,3 ± 0,74 (22,41-35,09); Longitud del Píleo (PL): 14,92 ± 0,39 (1,07-17,52); Anchura del Píleo (PW): 6,95 ± 0,15 (5,46-8,26); Longitud de la escama parietal (PaL): 5,22 ± 0,17 (3,39-6,44); Diámetro de la escama masetérica (DM): 2,18 ± 0,08 (1,46-2,87); Diámetro de la escama timpánica (DT): 2,01 ± 0,07 (1,13-2,83); Anchura de la escama anal (AW): 4,03 ± 0,12 (2,7-5,08); Altura o longitud de la escama anal (AL): 2,11 ± 0,07 (1,5-2,68); Número de gránulos supraciliares en el lado derecho (GrS r): 10,12 ± 0,34 (5-13); Número de gránulos supraciliares en el lado izquierdo (GrS l): 9,79 ± 0,38 (6-14); Gularia (GUL): 23,66 ± 0,31 (21-26); Collaria (COLL): 11,33 ± 0,26 (9-13); Dorsalia (DORS): 53,54 ± 0,72 (47-59); Ventralia (VENT): 26,37 ± 0,26 (24-29); Número de poros femorales en el lado derecho (FEM r):17,62 ± 0.25 (16-20); Número de poros femorales del lado izquierdo (FEM l): 17,54 ± 0,25 (15-20); Lamellas subdigitales (LAM): 25,25 ± 0,36 (23-30); Escamas circumanales (CircA): 6,58 ± 0,19 (5-8). Contactos entre placas: Proporción de contactos Rostral-internasal (R-I): 0,66 ± 0.17 (0-2); Proporción de contactos entre la Postocular y la Parietal (Po-Pa): 0,04 ± 0,02 (0-1); Proporción de contactos entre la Supranasal y la Loreal anterior (Sn-Lor): 0,18 ± 0.07 (0-1). Caracteres de coloración: Número de ocelos azules en un costado (BO): 2,79 ± 0.49 (0-11); Número de hileras de escamas a cada lado con puntuación ventral (PV): 1,75 ± 0,12 (1-3). Indices biométricos (ver abreviaturas en los caracteres lineales): FLL/SVL: 34,06 ± 0,004 (27,64-37,16); HLL/SVL: 49,83 ± 0,004 (45,25-53,46); PL/PW: 214,28 ± 0,02 (195,97-232,43); DM/PaL: 41,89 ± 0,01 (31,34-54,45); DT/PaL: 39,04 ± 0,01 (28,82-53,09); AL/AW: 52,44 ± 0,01 (43,60-64,11); AS/SVL: 477 ± 0,07 (404,31-545,5). Hembras: Basado en 26 ejemplares, se da la media ± desviación estandar (mínimo-máximo) (Arribas et al., 2006). Medidas: Longitud corporal (SVL): 64,19 ± 1,94 (48,25-84,42); Longitud de los miembros anteriores (FLL): 19,42 ± 0,42 (15,18-24,05); Longitud de los miembros posteriores (HLL): 27,37 ± 0,53 (22,75-32,98); Longitud del Píleo (PL): 13,52 ± 0,28 (11,27-16,65); Anchura del Pileo (PW): 6,62 ± 0,12 (5,6-7,9); Longitud de la escama parietal (PaL): 4,50 ± 0,11 (3,7-5,56); Diámetro de la escama masetérica (DM): 1,77 ± 0,08 (0,86-2,58); Diámetro de la escama timpánica (DT): 1,93 ± 0,06 (1,36-2,63); Anchura de la escama anal (AW): 3,72 ± 0,15 (2,15-5,28); Altura o longitud de la escama anal (AL): 2,18 ± 0,08 (1,52-3,29); Número de gránulos supraciliares en el lado derecho (GrS r): 9,65 ± 0,40 (6-13); Número de gránulos supraciliares en el lado izquierdo (GrS l): 9,53 ± 0,33 (5-12); Gularia (GUL): 23,88 ± 0,43 (16-29); Collaria (COLL): 11,23 ± 0,27 (9-14); Dorsalia (DORS): 53,11 ± 0,66 (48-61); Ventralia (VENT): 30,65 ± 0,24 (28-33); Número de poros femorales en el lado derecho (FEM r): 17,38 ± 0,34 (14-21); Número de poros femorales del lado izquierdo (FEM l): 17,23 ± 0,38 (13-21); Lamellas subdigitales (LAM): 25,34 ± 0,40 (22-30); Escamas circumanales (CircA): 6,23 ± 0,13 (5-8). Contactos entre placas: Proporción de contactos entre la escama Rostral y la internasal (R-I): 1,15 ± 0,15 (0-2); Proporción de contactos entre la Postocular y la Parietal (Po-Pa): 0,11 ± 0,05 (0-1); Proporción de contactos entre la Supranasal y la primera Loreal (Sn-Lor): 0,29 ± 0,08 (0-1). Caracteres de coloración: Número de ocelos azules en un costado (BO): 1,96 ± 0,24 (0-6); Número de hileras de escamas a cada lado con puntuación ventral (PV): 1,03 ± 0,12 (0-3). Indices biométricos (ver abreviaturas en los caracteres lineales): FLL/SVL: 30,49 ± 0,004 (26,75-34,62); HLL/SVL: 43,02 ± 0,005 (37,06-47,82); PL/PW: 203,97 ± 0,01 (196,44-219,37); DM/PaL: 39,27 ± 0,01(19,50-50); DT/PaL: 42,94 ± 0,01(35,07-59,52); AL/AW: 59,39 ± 0,01 (48,12-81,86); AS/SVL: 442,40 ± 0,07(366,66-529,88). Respecto a algunos caracteres morfológicos singulares, de diecinueve machos, quince tenían las prefrontales en contacto, como es típico y habitual en los lacértidos, mientras que dos ejemplares presentaban una escama supernumeraria entre ellas, y otros dos presentaban estas escamas separadas permitiendo el contacto entre las placas internasal y frontal. De veintiseis hembras, veintitrés tenían la disposición típica de las prefrontales, en pleno contacto, mientras que tres tenían una escama supernumeraria entre ellas. Nueve ejemplares de un total de cincuentaicinco (incluyendo ambos sexos) tienen dos masetéricas en vez de una. Coloración Coloración de los machos (periodo de celo, Sierra de la Cabrera): Tracto dorsal de verde a gris verdoso, tirando a verde grisáceo en el área pélvica y las patas posteriores. Ocelos axilares vivamente azules, muy numerosos, muchas veces de dos a siete, pero en ocasiones hasta once. Banda vertebral irregular (más o menos alineada en dos hileras yuxtapuestas de puntos) bien desarrollada y finamente disgregada, conectando con las bandas temporales (=costales), lo que da a los animales un aspecto general muy reticulado. Esta banda conecta rápida y progresivamente con las bandas costales a medida que el individuo crece. Los ocelos azules son reflectantes en ultravioleta y están presentes tanto en I. monticola e I. martinezricai, como en I. galani. En esta última especie, no obstante, los ocelos son más numerosos (y reflectantes en UV) mientras que están ausentes en Podarcis bocagei, sintópica con la lagartija leonesa en numerosas localidades. El número y tamaño de ocelos azules se incrementa con la edad (Arribas et al., 2006). Hembras (periodo de celo): No descrito. Machos (fuera del periodo de celo, de Sierra Segundera, Trevinca y Teleno): Color del fondo del tracto dorsal de grisáceo-amarillento a amarillo grisáceo, sólo ligeramente más oscuro cerca de la muda. Los ejemplares a medio crecer (especialmente los que viven sobre pizarras en el Teleno y Trevinca) tienen el dorso marronoso y frecuentemente bastante oscuro. Banda vertebral con manchas pequeñas o medianas e irregulares, incluso alargadas y vermiculadas, que están apenas alineadas en dos hileras de manchas yuxtapuestas pero distingibles. Estas manchas irregulares tienden a fusionarse con la edad. Bandas temporales (=costales) reticuladas, con el borde superior aserrado y unida por la parte inferior con las líneas laterales inferiores, quedando una hilera de puntos más claros entre ambas que es la prolongación de los ocelos axilares azules. Vientre verde pastel, y blanco verdoso hacia los límites del area gular. Los ocelos de los costados (ocelos axilares) son azules. Usualmente sólo las dos hileras de escamas más externas de cada lado del vientre presentan manchas oscuras bien visibles. Estas aparecen más centradas dentro de la escama en los machos jóvenes, pero son mayores y están conectadas al reborde anterior en los animales adultos. Existen manchas negras más escasas en las placas submaxilares y especialmente hacia los lados de la garganta (Arribas et al., 2006). Hembras (fuera del periodo de celo, de Sierra Segundera, Trevinca y Teleno): Dorso de color verde pálido a verde grisáceo en los adultos. Diseño mucho menos manchado que en los machos y con un grado de desarrollo no correlacionado con la edad (las hembras viejas pueden tener o carecer del moderadamente desarrollado diseño dorsal). Usualmente un moteado irregular o vermiculado en el centro del dorso, que puede extenderse a todo lo ancho del tracto dorsal o estar claramente alineado en dos hileras. Bandas temporales no reticuladas sino uniformes, con sus áreas marginales (especialmente el reborde superior) más oscuras (prácticamente negras) y el área interior de las bandas más clara (marrón). Borde superior de esta banda temporal (=costal) también aserrado, encerrando puntos más claros, más visibles en los animales jóvenes, pero también en los adultos. Las hileras de puntos claros en las partes inferiores de los costados menos marcadas, así como la línea lateral inferior, que es borrosa y raramente aparece en forma de manchas. Vientre verde amarillento. Ocelos azules como en los machos pero menos abundantes y más pequeños. Contrariamente a los machos, el moteado negro aparece usualmente en el centro (o en las partes poteriores) de las escamas ventrales de las hileras de escamas ventrales más externas, y frecuentemente está casi borrado. Más raramente existen pequeños puntos en la segunda hilera más externa de ventrales (Arribas et al., 2006).
Figura 2. Hembra de Iberolacerta galani. © O. Arribas. Crías (de Sierra Segundera y Teleno): Dorso con color de fondo amarillento grisáceo. Tracto dorsal finamente moteado con manchas irregulares en el area vertebral, que pueden ser muy borrosas y apenas distinguibles. Bandas temporales ligeramente reticuladas en las crías macho y más uniformes en las crías hembra (existe un cierto dimorfismo sexual desde el nacimiento). Vientre sin pigmento de color, con las ventrales más externas bien moteadas, pero con la segunda hilera (las intermedias) y la tercera (la interna) menos marcadas. Cola azul (Arribas et al., 2006).
Figura 3. Recién nacido de Iberolacerta galani. © O. Arribas.
Cariotipo Los ejemplares estudiados de Sanabria (macho y hembra) y de Teleno (hembra) muestran un cariotipo de 2n=36 macrocromosomas acrocéntricos gradualmente decrecientes en tamaño y con los organizadores nucleolares (NOR) en posición telomérica en un par de cromosomas grandes (tipo L) que podría tentativamente asimilarse al quinto par. La técnica del bandeo-C evidencia la presencia de cromosomas sexuales del tipo ZW y heteromórficos. El cromosoma W es tan grande como los autosomas del par 10 o 11 y completamente teñido con CMA3 y su heterocromatina teñida positivamente con DAPI. El cromosoma Z es tan grande como los cromosomas del sexto par, y difiere de los autosomas en que muestran gruesas bandas C teloméricas, teñidas con CMA3. Los autosomas muestran aparentemente bandas C centroméricas, teñidas con CMA3 y DAPI; así como bandas ligeras CMA3 y bandas C teloméricas. El cariotipo de I. galani se distingue de los cariotipos de I. martinezricai e I. cyreni por la posición y localización de los NORs (teloméricos y en un cromosoma grande en I. galani, e intersticiales y en un cromosoma medio-grande en I. martinezricai e I. cyreni). Difiere del cariotipo de I. monticola por la presencia en I. galani de cromosomas sexuales diferenciados, Z y W, ambos heteromórficos y heterocromáticos (homomórficos y eucromáticos en I. monticola). Difiere de I. horvathi (que también presenta cromosoma sexual W parcialmente heterocromático y NORs en un cromosoma grande, de tipo L) por el hecho de que en I. galani W está totalmente (no parcialmente) heterocromático y principalmente por la presencia de un cromosoma sexual Z diferenciado, único entre los Iberolacerta al tener una banda de heterocromatina CMA3 positiva y peritelomérica (Arribas et al., 2006). Osteología Se han estudiado dos ejemplares (macho y hembra) de Sanabria (Zamora) y una hembra del Macizo del Teleno (León). Todos ellos presentan 7 dientes premaxilares y un proceso nasal con lados sinuosos e irregulares, más o menos en forma de hoja (espatuliforme) o de punta de lanza (más claro en el macho estudiado). Huesos nasales relativamente cortos. Dieciseis o diecisiete dientes (posiciones dentales) en el maxilar, y de dieciocho a diecinueve en el dentario, dos tercios de ellos más o menos bicúspides, y el resto monocúspides. Sutura maxillo-yugal (margo ocularis) suave, no escalonada. Postorbital y postfrontal separados y subiguales en longitud. Proceso anteromedial del postorbital y anterodistal del postfrontal ambos presentes. El escamoso es bastante rectilíneo en comparación con otras Iberolacerta y en contacto con el postocular a lo largo de cerca de un tercio de la longitud de este último. No hay costillas asociadas a la tercera vértebra presacra. Formula costal esternal-xifisternal (3 + 2), y fontanela esternal prácticamente redonda. Clavículas de forma variable (abiertas-marginadas- en la hembra de Sanabria, y cerradas -emarginadas- en el macho de Sanabria y la hembra del Teleno estudiados). Interclavícula cruciforme con ramas laterales muy esbeltas. Estas ramas laterales tienen longitudes muy similares a la rama posterior. Relación entre las ramas anterior y posterior desde 0,40 (hembras) a 0,42 (macho). El ejemplar macho posee 26 vértebras presacrales y las hembras 28-29, las últimas seis asociadas a vértebras cortas. La hembra del Teleno presenta un conteo de 29 vértebras en el lado izquierdo y 28 en el derecho, debido que la vértebra presacra nº 29 poseee una costilla hacia un lado y un proceso que forma parte del sacro hacia el otro. La quinta vértebra preautotómica es del tipo-A de Arnold (1973). Osteológicamente, I. galani es muy similar a las otras especies del “grupo ibérico”. No obstante, difiere en la forma del hueso escamoso, que es claramente curvado en todas las otras Iberolacerta (incluyendo las pirenaicas y la latealpina I. horvathi) pero bastante rectilíneo en I. galani. Además, en el “grupo ibérico”, I. cyreni tiene usualmente huesos nasales más largos y nueve dientes premaxilares (siete en las I. galani estudiadas); I. monticola tiene un proceso nasal espatuliforme o alargado y de lados más o menos paralelos (sólo raramente en forma de punta de flecha, más frecuente en Galicia y Estrela, e igualmente presente en I. galani); I. martinezricai tiene nueve dientes premaxilares, un proceso nasal elongado (de lados paralelos, no en punta de flecha como ocurre en I. galani), y frecuentemente siete vértebras presacrales cortas y un fino proceso medial de forma triangular en el escamoso, al menos en algunos especímenes (Arribas et al., 2006).
Variación geográfica Los ejemplares que habitan sobre pizarras (p., ej. Teleno) son notablemente más oscuros y menos reticulados que los ejemplares de zonas graníticas (Sanabria). Los datos sobre los valores conocidos de folidosis y biometría de las distintas poblaciones se encuentran en Arribas et al. (2006). Existen heterozigotos en la posición 22 del fragmento de 342 bp del gen c-mos secuenciado en Arribas et al. (2006). En esta posición, todas las muestras estudiadas del grupo ibérico tienen una C, con la excepción de todas las muestras de I. monticola, que tienen una T y algunas muestras de I. galani que tienen tanto una T (muestras de Peña Trevinca, del Puerto de los Portinillos y del Puerto El Morredero) o son heterozigotos (C/T) para esta posición (muestras de Peña Trevinca, de Laguna del Sotillo, de Laguna de los Peces, y del Puerto El Morredero). Todas las demás muestras de I. galani tienen una C en la posición 22 (muestras de la Laguna de los Peces). No existe un patrón geográfico en esta variabilidad genética (Arribas et al., 2006). Se han descrito 14 loci microsatélites en I. monticola que también son polimórficos en I. galani (Remón et al., 2008).1
Referencias Arnold, E. N. (1973). Relationships of the Palaearctic lizards assigned to the genera Lacerta, Algyroides and Psammodromus (Reptilia: Lacertidae). Bulletin of the British Museum (Natural History), 25: 292-356. Arribas, O. (2014). Iberolacerta (Iberolacerta) galani Arribas, Carranza y Odierna, 2006. Pp. 398-409. En: Salvador, A. (Coordinador). Reptiles, 2ª edición revisada y aumentada. Fauna Ibérica, vol. 10. Ramos, M. A. et al. (Eds.). Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas, Madrid. 1367 pp. Arribas, O., Carranza, S. (2004). Morphological and genetic evidence of the full species status of Iberolacerta martinezricai (Arribas, 1996). Zootaxa, 634: 1-24. Arribas, O., Carranza, S., Odierna, G. (2006). Description of a new endemic species of mountain lizard from Northwestern Spain: Iberolacerta galani sp. nov. (Squamata: Lacertidae). Zootaxa, 2240: 1-55. Carranza, S., Arnold, E. N., Amat, F. (2004). DNA phylogeny of Lacerta (Iberolacerta) and other lacertine lizards (Reptilia: Lacertidae): did competition cause long-term mountain restriction? Systematics and Biodiversity, 2: 57-77. Holdhaus, K. (1902). Beitrage zur Koleopteren-Geographie. Muench. Koleopterol. Z., 1: 255-262. Holdhaus, K. (1906). Die verbreitung der Coleopteren in den mitteleuropaischen Hochgebirgen. Verh. Zool. Bot. Ges. Wien, 56: 629-641. Holdhaus, K. (1954). Die Spuren der Eiszeit in der Tierwelt Europas. Abh. Zool. Bot. Ges. Wien, 18: 1-493. Mayer, W., Arribas, O. (1996). Allozyme differentiation and relationships among the Iberian-Pyrenean Mountain lizards (Squamata: Sauria: Lacertidae). Herpetozoa, 9: 57-61. Remón, N., Vila, M., Galán, P., Naveira, H. (2008). Isolation and characterization of polymorphic microsatellite markers in Iberolacerta monticola, and cross-species amplification in Iberolacerta galani and Zootoca vivipara. Molecular Ecology Resources, 8 (6): 1351-1353.
Oscar Arribas Salvador Carranza Fecha de publicación: 21-03-2007 Otras contribuciones: 1. Alfredo Salvador. 25-08-2009; 2. Alfredo Salvador. 14-07-2015 Arribas, O., Carranza, S. (2015). Lagartija leonesa – Iberolacerta galani. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Marco, A. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|
|
|
|
|
|