|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Pyrenean desman, identification, size, variation.
Nombres comunes En la mayor parte de los idiomas Galemys pyrenaicus es conocido como desmán de los Pirineos a causa de que su descripción científica se hizo a partir de un ejemplar capturado en Tarbes, una localidad del los Pirineos franceses, por Etienne Geoffroy (1811) y hasta bien entrado el siglo XX la mayor parte de los estudios y por tanto del conocimiento de la especie se hicieron en Francia, país en el que la especie está presente solamente en esta cordillera y en sus inmediaciones, lo que explica el adjetivo “pirenaico” que habitualmente le ha acompañado. Su descubrimiento posterior en Sistema Central (Graells, 1852), cerca del El Escorial, y posteriormente en Asturias (Pastor, 1859), Galicia (López Seoane, 1863) y Portugal (Trutat, 1891) amplió notablemente el área de distribución de la especie, por lo que, teniendo en cuenta que la mayor parte de su distribución se encuentra fuera de la cadena pirenaica resulta mucho más adecuado el nombre de desmán ibérico. Graells (1897) recoge diversos nombres vernáculos en Castilla, como almizclera, rata almizcleña; en Cataluña, Rata d’aigua y almesquera y en Galicia, Aguaneira y rato d’almiscle. Inglés: Pyrenean desman. Francés: Desman des Pyrénées. Castellano: Desmán de los Pirineos, Desmán ibérico, Almizclera. Portugués: Toupeira-de-água. Catalán: Almesquera. Euskera: Muturluze iberiarra. Gallego: Rato de almizcre.
Nomenclatura El primer desmanino en ser descrito por la ciencia fue el desmán ruso, habitante de las cuencas de los Ríos Don, Volga y Ural. Linné (1758) describe de oídas un Castor moschatus, que vive en las aguas rusas y en América. Aunque esta descripción representa una confusión parcial con la rata almizclera, al ser ambos mamíferos pequeños, de hábitos semiacuáticos, utilizados en peletería, parte del error es corregido por el propio Linné siete años más tarde dando a la segunda el nombre de Castor zibethicus. Posteriormente Cuvier (1800) crea un nuevo género, Mygale (de xum, ratón y hlag mustélido), pero como este nombre ya había sido empleado para un género de insectos, hubo que utilizar variantes del mismo, como Myogalea y Myogale, que fue el género con el que fue conocido durante todo el siglo XIX. Etienne Geoffroy (1811) describió una nueva especie sobre un ejemplar procedente de Tarbes (Hauts-Pyrenées), con el nombre de Mygale pyrenaica. Kaup describió un nuevo género, Galemys, para los desmanes de Europa Occidental, mucho más pequeños que los rusos y Miller (1912), en su revisión taxonómica de los mamíferos europeos, estableció la nomenclatura que actualmente se utiliza, denominando Desmana moschata a la especie rusa y Galemys pyrenaicus a la ibero-francesa.
Filogenia Los datos moleculares muestran que los linajes de los topos euroasiáticos y los desmanes se separan hace 37 millones de años, es decir, en el ecuador del período Terciario, coincidiendo con una pequeña edad de hielo, preludio del mayor enfriamiento del final del Eoceno (Douady y Douzery, 2003). Rümke (1985) ha descrito la filogenia de los desmaninos a partir de los fósiles. Este grupo es conocido en Europa y Asia desde el Mioceno final a partir del género Dibolia, en el que se produce la separación del linaje occidental del oriental, que probablemente conduce a los géneros actuales. Generalmente se considera que la subfamilia es todavía más antigua y la retrotraen al inicio del Mioceno con el género Mygalea (Ziegler, 2003) en Centroeuropa, lo que supone que ya existían hace unos 24 millones de años, edad más aproximada a la divergencia señalada por los relojes moleculares y que probablemente señala el abandono de la línea excavadora por una creciente especialización acuática. Uno de los caracteres diagnósticos de la subfamilia es la forma del húmero, no adaptada a la actividad excavadora, de modo que aparentemente podían haber sido desde el principio nadadores. Galemys pyrenaicus es el grupo hermano de Desmana moschata, lo que confirma la monofilia de la subfamilia Desmaninae (Cabria et al., 2006). Los fósiles del género Galemys (G. sulimskii de Polonia) se conocen desde el final de Plioceno y se encuentran fósiles de G. kormosi repartidos por casi toda Europa, desde Polonia a España y desde Hungría a Inglaterra en el Pleistoceno Inicial, de forma que a partir de este momento se produce una contracción del área de distribución hacia el suroeste para quedar reducida al ámbito Ibero-Pirenaico de la especie actual (Harrison et al., 1988). La explicación de esta contracción probablemente no haya que buscarla en la exclusión provocada por la irrupción de Neomys en el continente europeo como señalan Harrison et al. (1988), ya que en toda la extensión actual de Galemys pyrenaicus convive en simpatría con Neomys spp., sino probablemente en una especialización creciente a las aguas de montaña de los desmaninos de menor tamaño, a causa de lo cual encontraron refugio en las montañas ibéricas que no sufrieron procesos de glaciación tan intensos como en otras montañas europeas. Aunque parece haber cierta confusión en la determinación y denominación de algunas formas fósiles en décadas pasadas (Palmeirim y Hoffmann, 1983), Galemys pyrenaicus aparece sólo en el entorno ibérico y se encuentra ya en el Pleistoceno Inferior (Arribas, 2004; Sesé y Sevilla, 1996).
Descripción Es un animal inconfundible, su trompa aplastada y desnuda, destaca de un cuerpo rechoncho, con una gruesa cola escamosa, de sección redondeada, pero comprimida lateralmente en su extremo (Figura 1). Nos basaremos en la descripción realizada por Cabrera (1914) para la especie, dado que se trata de un género monoespecífico sin ninguna similitud estrecha con ninguna otra especie de la fauna ibérica.
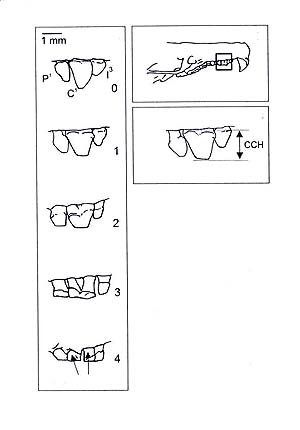
Figura 1. Desmán ibérico. © P. García-Rovés Caracteres externos Trompa bastante larga, muy aplastada, con las narices abiertas transversalmente en la cara dorsal de su extremo, desnuda, pero franjeada en los lados por abundantes y largas vibrisas. Ojos pequeños, prácticamente invisibles bajo el pelo. Cuerpo rechoncho, que en estado de reposo parece redondeado, mientras que cuando nada se alarga considerablemente. Hay que tener en cuenta que en muchas guías de campo el cuerpo aparece excesivamente alargado, probablemente debido a haber sido dibujado a partir de ejemplares naturalizados, que no recuerdan en absoluto al aspecto del animal vivo. Extremidades anteriores pequeñas, con membrana interdigital corta. Pies posteriores grandes, largos, encorvados hacia adentro, con amplia membrana interdigital, cubiertos de piel escamosa y con una franja de pelos largos, fuertes y compactos. Cola muy larga, cilíndrica, comprimida lateralmente hacia la punta y revestida de pelo duro, bastante escaso para dejar entrever la piel escamosa. Bajo la base de la cola se abre una glándula que segrega un humor almizclado. Mamas 1.1-3.3. Color pardo oscuro, variando del sepia al pardo claro, con brillantes reflejos metálicos plateados o bronceados, que se acentúan cuando el animal está sumergido en el agua. La trompa y las extremidades tienen la piel negruzco, mientras los pelos que hay en estas artes son plateados o de un color de ante muy lustroso. Región ventral mucho más pálida que la dorsal, de un gris reluciente cuyo matiz varía mucho, según los ejemplares. Cola de color carne lívido con los pelos blancuzcos o amarillentos. El pelo del desmán tiene especial significación para la confirmación en la identificación de los excrementos, ya que son accidentalmente ingeridos como consecuencia del grooming (operación de autolimpieza en la que se pasa la lengua por el pelaje). Los mamíferos tienen dos tipos de pelos: la borra, pelo lanoso que forma el aislante térmico y la jarra, pelo más largo y rígido, visible al exterior, que protege externamente la borra evitando que se moje. El desmán tiene la particularidad de tener dos tipos de jarra, una de ellas fácilmente identificable porque carece de médula, está doblado hacia adentro en su mitad distal y a continuación presenta un aplanamiento apical en forma de punta de lanza, lo que constituye un carácter diferencial de este tipo de jarra, llamada granne por Poduschka y Richard (1985). Cráneo Cráneo con el rostro muy estrecho y prolongado, relativamente parecido al de los topos. Caja cerebral ensanchada y truncada posteriormente, presentando, cuando se la mira por encima, la forma de un triángulo isósceles cuya base corresponde a la región occipital. Un agujero preorbitario bastante grande delante de cada arco cigomático. Mandíbula con una apófisis coronoide muy elevada. La forma de la mandíbula se correlaciona con su tamaño y una vez que los jóvenes abandonan el nido, la geometría mandibular no muestra variación significativa, siendo independiente de factores geográficos (Ventura y López-Fuster, 2010)1. Fórmula dentaria 3.1.4.3/3.1.4.3. Incisivos superiores centrales grandes, puntiagudos, en forma de pirámide triangular, de bordes cortantes; mientras que el segundo y tercero son sumamente pequeños. El canino superior es sólo un poco mayor que el incisivo que le precede y aproximadamente igual a los tres primeros premolares. Molares parecidos a los de Talpa, pero el m1 y el m2 con nueve cúspides, mientras que en Talpa sólo hay siete u ocho (falta el protocónulo). El canino inferior apenas se diferencia del incisivo inmediato. El cariotipo es de 2n = 24 cromosomas, aunque hay diferentes interpretaciones en relación con el número de brazos que Peyre (1957a) cifra entre 65 y 67, a causa de la cortedad de los brazos del par 14, que Ramalhinho (1990) no considera, centrándolo en 66. Identificación genética Se ha propuesto un método PCR-RFLP para identificar genéticamente el desmán en muestras de ADN procedente de heces (Gillet et al., 2015)2. Sexado El desmán no presenta un evidente dimorfismo sexual, como es la regla general entre los tálpidos. Resulta difícil sexar los animales en vivo, dado que ambos sexos tienen un órgano peniforme, los machos tienen los testículos intraabdominales y las hembras tienen la vagina abierta durante un breve período de tiempo. Los desmaninos, como otros tálpidos presentan un arco pelviano muy fino, que une las dos extremidades isquiopubianas, que puede apreciarse por palpación (Peyre, 1957b) que ha sido aprovechado en varios estudios para identificar el sexo de los desmanes capturados en vivo. En la hembra el arco es de consistencia dura y evidente cuando es inmadura, dado que el arco es cartilaginoso, comienza a presentar una ligera hendidura central en los prepúberes, que se va ampliando, y en las hembras adultas se manifiesta un amplio espacio (unos 5 mm ) tras sufrir una osteolisis completa que convierte el arco en un ligamento conjuntivo. En los machos, el arco pelviano está siempre duro, ya que partiendo también de un arco cartilaginoso acaba por osificarse completamente en los adultos. Durante la época de celo las hembras pueden distinguirse gracias al orificio vaginal abierto y depigmentado (Richard, 1986) González Esteban et al. (2003) han identificado el sexo por observación de los genitales externos aplicando una ligera presión a la papila urinaria, por ambos lados y hacia abajo, con el fin de evaginarlos. Los machos entonces muestran un glande en forma de cono, más o menos alargado, que en los adultos es más ancho en su base. En las hembras se observa una papila urinaria, larga y estrecha, con una especie de muesca subterminal. Se ha establecido un protocolo de determinación del sexo en muestras no invasivas basado en la secuenciación parcial de fragmentos de los genes DBX y DBY (Vidal et al., 2010)1. Determinación de la edad La masa corporal puede considerarse a priori el modo más sencillo para distinguir los animales juveniles del año de los adultos en los estudios de campo. Stone (1987) ha considerado animales juveniles aquellos que pesaban menos de 45 g, sin embargo González-Esteban et al. (2002) consideran que los animales jóvenes se encuentran en la población con pesos similares a los adultos y no encuentran correlación entre la edad y la masa corporal de los animales, al menos a partir de uno cuatro meses de edad. Richard (1976) fue el primero en establecer un criterio de desgaste de los dientes. Utilizó un índice que representa la suma del desgaste de las coronas de los caninos y de los tres primeros premolares de cada hemimaxilar y de los terceros incisivos, caninos y tres primeros premolares de cada hemimandíbula, en total 18 medidas. González-Esteban et al (2002) establecen unos criterios cuantitativos y semicuantitativos más sencillos basados únicamente en el desgaste del canino superior y encuentran que el recuento de los anillos de cemento del canino o del primer premolar permiten contar igualmente el número de años cumplidos por los ejemplares. El criterio cuantitativo se basa en el valor relativo de la altura de la corona del canino superior (HCC) en relación con la longitud condilobasal del cráneo (LCB): D = (HCC/CLB) x 100 El criterio semicuantitativo establece las siguientes clases (Tabla 1) (Fig. 2).
Tabla 1. Criterios semicuantitativos de determinación de la edad. Según González-Esteban et al. (2002).
Figura 2. Desgaste dental de los desmanes ibéricos. Derecha arriba: Rostro de un cráneo de desmán en vista lateral. El cuadrado muestra el fragmento representado debajo, indicando la altura de la corona del canino superior CCH. Izquierda: Forma del canino superior (en vista labial) para cada una de las cinco clases establecidas: P1 – primer molar superior. C1 – primer canino superior. I3 – tercer incisivo superior (Según González-Esteban et al., 2002).
Tamaño La longitud de cabeza y cuerpo varía entre 108 y 135 mm, la longitud de la cola entre 100 y 156 mm y la longitud del pié posterior entre 30 y 38 mm (Niethammer, 1970; Peyre, 1962). Ver medidas craneales en Tabla 2.
Tabla 2. Medidas craneales de Galemys pyrenaicus en mm (González Esteban et al., 1999). Acran: Anchura craneana; LSDS: Longitud de la serie dentaria superior; AIO: Anchura interorbitaria; LCB: Longitud condilobasal; Acor: Altura de la rama coronoide de la mandíbula; LM: Longitud de la mandíbula.
Masa corporal La masa corporal varía entre 54 y 80 g (Niethammer, 1970; Peyre, 1962; Juckwer, 1990). Ver Tabla 3 sobre variación geográfica de la masa corporal.
Variación geográfica Graells (1897) describió Myogalea rufula por la coloración parda con reflejos plateados o dorados. Las revisiones de Miller (1912) y Cabrera (1914) quitan validez taxonómica a la coloración, que consideran fruto por una variabilidad individual, pero mantienen la validez subespecífica Galemys pyrenaicus rufulus (Graells, 1897) a causa del mayor tamaño de los ejemplares que se encuentran desde Galicia hasta el Sistema Central, considerando la subespecie típica Galemys pyrenaicus pyrenaicus (E. Geoffroy, 1811) para los Pirineos y la Cordillera Cantábrica. Este criterio ha sido mantenido por Niethammer (1970) y Juckwer (1990) (Tabla 3), si bien relativizando las diferencias, que consideran escasas, teniendo en cuenta ejemplares procedentes de Navarra, Cantabria, León, La Rioja , Burgos y Ávila. González-Esteban et al. (1999), con abundante material de Galicia, Cordillera Cantábrica, Sistema Ibérico Norte, pero con un solo ejemplar del Sistema Central (Sierra de Gredos) encuentran que ni la coloración ni las medidas apoyan los criterios subespecíficos clásicos, ya que los ejemplares orientales, procedentes de Pirineos, País Vasco y Sistema ibérico Norte son de color pardo oscuro (negruzcos), mientras que los occidentales, de Asturias, León y Galicia son de color pardo claro (rojizos), mientras que el tamaño de los ejemplares pirenaicos es intermedio entre los de Galicia y el Sistema Ibérico que Miller y Cabrera habían asignado a rufulus.
Tabla 3. Variación geográfica de la masa corporal en poblaciones de Galemys pyrenaicus.
Por su parte López-Fuster et al. (2006), constatan que hay una variación de tamaño en toda su área de distribución. Consideran que los de Pirineos y Galicia son menores que los de la Cordillera Cantábrica y los de los sistemas Central e Ibérico, pero las relaciones canónicas de diversas medidas craneales diluyen a los ejemplares gallegos entre los representantes de las poblaciones no pirenaicas, por lo que no descartan de forma tan drástica las diferencias entre pyrenaicus y rufulus. Hay una fuerte estructura filogeográfica, influida por las glaciaciones del Pleistoceno (Igea et al., 20132), con cinco clados que se corresponden con Pirineos, Cordillera Cantábrica, Sistema Ibérico, Sistema Central y noroeste Ibérico (Galicia, norte de Portugal y Montes de León) (Querejeta et al., 2016)2.
Referencias Arribas, O. (2004). Fauna y paisaje de los Pirineos en la Era Glaciar. Lynx Ediciones-Fundació Territori i Paisatge, Barcelona. 540 pp. Cabrera, A. (1914). Fauna Ibérica. Mamíferos. Mus. Nacional Cien. Nat., Madrid. 441 pp. Cuvier, G. (1800). Leçons d'anatomie comparée. Tome I. Baudouin, Paris. 520 pp. Cabria, M. T., Rubines, J., Gómez-Moliner, B., Zardoya, R. (2006). On the phylogenetic position of a rare Iberian endemic mammal, the Pyrenean desman (Galemys pyrenaicus). Gene, 375: 1-13. Douady, C. J., Douzery, E. J. P. (2003). Molecular estimation of eulipotyphlan divergence times and the evolution of “Insectivora”. Mol. Phylogen. Evol., 28: 285-296. Geoffroy, E. (1811). Memoire sur les espéces des genres Musaraigne et Mygale. Ann. Mus. Hist. Nat., 17: 169-194. González-Esteban, J., Castién, E., Gosálbez, J. (1999). Morphological and colour variation in the Pirenean desman Galemys pyrenaicus (Geoffroy, 1811). Z. Säugetierkd., 64: 1-11. González-Esteban, J., Villate, I., Castién, E., Rey, I., Gosálbez, J. (2002). Age determination of Galemys pyrenaicus. Acta Theriologica, 47 (1): 107-112. González-Esteban, J., Villate, I., Castién, E. (2003). Sexual identification of Galemys pyrenaicus. Acta Theriologica, 48 (4): 571-573. Graells, M. de la P. (1852). Sección Zoológica. En: F. Luján. Memoria que comprende el resumen de los trabajos verificados en 1851 por las diferentes secciones encargadas de formar el mapa geológico de la provincia de Madrid y en general de todo el Reino. Madrid. Graells, M. de la P. (1897). Fauna Mastodológica Ibérica. Mem. Acad. Cien. Exact. Fis. Natur., 17: 1-806. Gillet, F., Cabria, M. T., Nemoz, M., Blanc, F., Fournier-Chambrillon, C., Sourp, E., Vial-Novella, C., Aulagnier, S., Michaux, J. R. (2015). PCR-RFLP identification of the endangered Pyrenean desman, Galemys pyrenaicus (Soricomorpha, Talpidae), based on faecal DNA. Mammalia, 79 (4): 473-477. Igea, J., Aymerich, P., Fernández-González, A., González-Esteban, J., Gómez, A., Alonso, R., Gosálbez, J., Castresana, J. (2013). Phylogeography and postglacial expansion of the endangered semi-aquatic mammal Galemys pyrenaicus. BMC Evolutionary Biology, 13: 1-19. Juckwer, E. A. (1990). Galemys pyrenaicus (Geoffroy, 1811) – Pyrenäen-Desman. Pp. 79-92. En: Niethammer, J., Krapp, F. (Eds.). Handbuch der Säugetiere Europas. Vol. 3/1. Akademische Verlag, Wiesbaden. Linné, C. (1758). Systema naturae. Tomus I. Imp. Laurentii Salvii, Holmiae. 824 pp. López-Fuster, M. J., García-Perea, R., Fernández-Salvador, R., Gisbert, J., Ventura, J. (2006). Craniometric variability of the Iberian desman, Galemys pyrenaicus (Mammalia: Erinaceomorpha: Talpinae). Folia Zool., 55 (1): 29-42. López Seoane, V. (1863). Fauna mastológica de Galicia. Imprenta Mauel Mirás, Santiago de Compostela. 544 pp. Miller, G. S. (191). Catalogue of The Mammals of Western Europe . British Mus. (Nat. Hist.), Londres. 1017 pp. Niethammer, J. (1970). Beobachtungen am Pyrenäen-Desman, Galemys pyrenaica. Bonn . Zool. Beitr., 21: 157-182. Palmeirim, J. M., Hoffmann, R. S. (1988). Galemys pyrenaicus. Mammalian Species, 207: 1-5. Pastor, P. (1859). Apuntes sobre la fauna asturiana. Imprenta de Benito González, Oviedo. 4 pp. Peyre, A. (1957a). La formule chromosomique du desman des Pyrénées Galemys pyrenaicus G. Bull. Soc. Zool. France, 82 (5-6): 434-437. Peyre, A. (1957b). Dimorphisme sexuel de la centure pelvienne d’un mammifère Insectivore, Galemys pyrenaicus G. C. R. Acad. Sci. Paris , 244: 118-120. Poduschka, W., Richard, B. (1985). Hair types in the fur of the Pyrenean desman (Galemys pyrenaicus) Geoffroy, 1811 (Insectivora: Talpidae: Desmaninae). Oesterr. Akad. Wiss. Math. Naturwiss. Sitzungsber.Abt. I, 194 (1-5): 39-44. Puisségur, C. (1935). Recherches sur le desman des Pyrénées. Bull. Soc. Hist. Nat. Toulouse, 67: 163-227. Querejeta, M., González-Esteban, J., Gómez, A., Fernández-González, A., Aymerich, P., Gosálbez, J., Escoda, L., Igea, J., Castresana, J. (2016). Genomic diversity and geographical structure of the Pyrenean desman. Conservation Genetics, 17 (6): 1333-1344. Ramalhinho, M. G. (1990). Chromosomes of Galemys pyrenaicus (E. Greoffroy, 1811) from Portugal. Arq. Mus. Bocage Nova Sér., 1 (40): 579-583. Richard, P. B., Vallete Vaillard, A. (1969). Le desman des Pyrénées (Galemys pyrenaicus): premiéres notes sur sa biologie. Terre Vie, 3: 225-245. Richard, P. B. (1976). Determination de l’age et de la longevité chez le desman des Pyrenées (Galemys pyrenaicus). Terre Vie, 30: 181-192. Rümke, C. G. (1985). A review of fossil and recent desmaninae (Talpidae, Insectivora). Utrecht Micropal. Bull. Sp. Publ., 4: 1-241. Sesé, C., Sevilla, P. (1996). Los micromamíferos del Cuaternario peninsular español: Cronoestratigrafía e implicaciones bioestratigráficas. Rev. Española Paleo., Nº Extra.: 278-287. Shinohara, A., Campbell, K. L., Suzuki, H. (2003). Molecular phylogenetic relationships of moles, shrew moles, and desmans from the new and old worlds. Mol. Phyl. Evol., 27: 247-258. Stone, R. D. (1987). The social ecology of the Pyrenean desman (Galemys pyrenaicus) (Insectivora: Talpidae), as revealed by radiotelemetry.J. Zool. Lond., 212: 117-129. Trutat, E. (1891). Essai sur l’Histoire naturelle du Desman des Pyrenées. Impr. Douladoure, Toulouse. 107 pp. Ventura, J., López-Fuster, M. J. (2010). Geometric morphometrics of the mandible in the Iberian desman, Galemys pyrenaicus (Mammalia: Soricomorpha): Is there a significant variation in form during post-weaning life? Mammalian Biology, 75 (3): 191-197. Vidal, O., Pérez-Serra, A., Pla, C. (2010). A sex determination protocol for the Iberian desman (Galemys pyrenaicus) based on a three primer amplification of DBX and DBY fragments with non-invasive samples. Conservation Genetics, 11 (3): 1185-1187. Ziegler, R. (2003). Moles (Talpidae) from the late Middle Miocene of South Germany. Acta Paleontologica Polonica, 48 (4): 617-648.
Carlos Nores Fecha de publicación: 5-07-2007 Otras contribuciones: 1. Alfredo Salvador. 19-11-2012; 2. Alfredo Salvador. 18-01-2017 Nores, C. (2017). Desmán ibérico – Galemys pyrenaicus. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Barja, I. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||