|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Eurasian Curlew, habitat, abundance, status, threats.
Hábitat A nivel europeo es una limícola altamente adaptable en relación a la selección de zonas para nidificar, utilizando tanto medios naturales como otros resultantes de usos agrícolas o ganaderos (Cramp y Simmons, 1993; Dubois y Mahéo, 1986). En general prefiere áreas abiertas no perturbadas, evitando setos, postes y carreteras (Wilsonet al., 2004), así como también la proximidad de bordes de bosque (Valkama y Currie, 1999). Selecciona preferentemente áreas donde la superficie ocupada por pastizales sea mayor del 50% (Atkinson et al., 2002) y con alturas del estrato herbáceo comprendidas entre 10 y 45 cm (Durant et al., 2008). Para ubicar el nido parece evitar campos con vegetación muy baja (Valkama y Currie, 1999). La población nidificante española de la comarca de Terra Chá (Lugo), ocupa prados de siega, utilizados fundamentalmente como zonas de alimentación (Figura 1), y brezales-tojales de poca altura y con espacios de suelo descubierto, próximos a zonas de turbera, rodeados por prados de siega y campos de cultivo (Domínguez y Vidal, 2009). La actual zona de ubicación de nidos es un área de landa y matorral, cercana a 300 ha, en la que predomina el matorral de bajo porte (< 30 cm) de Erica sp. y Calluna alternando con zonas de Ulex cerrado y de altura elevada (1-1,5 m) (Figura 2). En la tipología CORINE esta zona se define como de landas y matorrales templado-oceánicos. También se encuentran pequeñas extensiones de turbera, de notable interés por ser utilizadas por adultos y pollos no volantones a medida que progresa la primavera y se evidencia el estiaje. Las características del matorral, altura y cobertura, son cambiantes, debido fundamentalmente a incendios y desbroces (Domínguez y Vidal, 2009). En la comarca de A Limia (Ourense), donde nidificó de forma regular hasta la década de 1960 y esporádicamente con posterioridad, las aves utilizaron principalmente labradíos y barbechos húmedos o parcialmente inundados (casi 2/3 de todas las observaciones), aunque también se registraron en pastizales higrófilos y vegas encharcadas (Villarino et al., 2002).
Figura 1. Prados de siega y zona de matorral bajo utilizados en la búsqueda de alimento y ubicación de nidos. (C) J. Domínguez.
Figura 2. Vista de la zona de ubicación de nidos de Zarapito real en Terra Chá (Lugo). (C) J. Domínguez.
En Asturias, donde hay constancia de su reproducción hasta 1977, utilizó para nidificar campos de ladera en colinas y terrenos ganados a la marisma, inundables con ocasión de fuertes mareas y temporales (Noval y Cortés, 2000). Las poblaciones invernantes utilizan a menudo los pastos litorales, especialmente durante la marea alta (Navedo et al., 2013). En periodo otoñal e invernal la población ibérica se nutre de efectivos, procedentes de países europeos, que se distribuyen mayoritariamente por humedales costeros (Catry et al., 2010; González y Pérez-Aranda, 2011) y con presencia regular, aunque escasa, en localidades continentales (Jubete, 2005; González y Pérez-Aranda, 2011). Los principales humedales de importancia para la invernada se localizan en la fachada cántabro-atlántica, destacando las marismas de Santoña, las rías de Ortigueira y Arousa y la bahía de Cádiz (Domínguez, 1997; González y Pérez-Aranda, 2011). En el periodo 1990-2009 la población media invernante en España fue de 4.148 ejemplares, con una tendencia de incremento moderado (González y Pérez-Aranda, 2011), mientras que en Portugal la población media en el trienio 2005-2007 fue de 2.280 ejemplares (Catry et al., 2010).
Abundancia La densidad de parejas nidificantes es dependiente del tipo de medio ocupado, aunque no existe una relación precisa entre la densidad de parejas y la abundancia de alimento. En Europa se han mencionado valores comprendidos entre 0,1 y 6 parejas/10 ha (Tabla 1), si bien los criterios empleados para delimitar el área ocupada dificultan en cierta medida su interpretación. La densidad de la población de Terra Chá, considerando exclusivamente el área de landa y matorral donde se han encontrado los nidos entre 2007 y 2012, se sitúa en valores bajos dentro del rango descrito para distintas zonas europeas.
Tabla 1. Densidad de parejas nidificantes de Zarapito real en diferentes zonas europeas. El asterisco indica que el dato corresponde a individuos/10 ha.
Tamaño poblacional La población europea se ha estimado comprendida entre 700.000-1.000.000 de ejemplares (Scott, 2009) y la de la Unión Europea entre 183.000 y 214.500 parejas (Thorup, 2006). Los mayores efectivos reproductores en la Unión Europea se localizan en Gran Bretaña y Finlandia, disminuyendo considerablemente su abundancia hacia el sur del continente (Thorup, 2006). En la península Ibérica sólo hay constancia de nidificación pasada y actual en España, ya que en Portugal no se han documentado episodios de cría (Tait, 1887, 1924; Catryet al., 2010), si bien Tait (1924) indicó la presencia de aves con comportamiento de cortejo y formación de pareja en marzo, aunque sin concretar la localidad de observación. La población reproductora española no se mencionó en catálogos de aves publicados en el siglo XIX y primeras décadas del XX, tanto de ámbito nacional (Reyes y Prosper, 1886; Arévalo Baca, 1887) como de Galicia (Ríos Naceyro, 1850; López Seoane, 1886; Iglesias, 1927). No obstante, a principios del siglo XX esta limícola fue citada por Rios Naceyro como residente en Galicia (Tait, 1924), lo que sugiere que el naturalista gallego quizás dispusiera de datos sobre su nidificación. Revisiones españolas de mediados del siglo XX trataron al Zarapito real como reproductor dudoso (Bernis, 1954) o con posibles intentos de reproducción muy esporádicos en el noroeste ibérico y marismas del Guadalquivir (Bernis, 1966). En obras de referencia sobre la avifauna europea se mencionó como nidificante en Asturias y, excepcionalmente, Andalucía (Glutz Von Blotzheimet al., 1986; Cramp y Simmons, 1993). Más recientemente, en el Atlas provisional de aves nidificantes de Galicia, que recogía información del periodo 1970-1979, no se citó su reproducción (López y Guitián, 1983), si bien la cobertura espacial de este primer atlas gallego fue muy incompleta. Sí fue mencionado como nidificante en el Atlas de Aves de Galicia correspondiente al periodo 1980-1985 (Bárcena y Domínguez, 1995) y en el primer Atlas de Aves de ámbito español, que recogió información del periodo 1975-1995 (De Juana, 1997). Las poblaciones actuales más cercanas a la de Galicia se encuentran en el sur de Francia y son de pequeño tamaño (Deceuninck, 2001; Issa y Boutin, 2010). La población española se localiza en la provincia de Lugo. En 2007 el censo fue de 3 parejas, presentes en el ayuntamiento de Castro de Rei (Domínguez y Vidal, 2009). Entre 2008 y 2011 la población fue similar a la de 2007, mientras que en 2012 se censaron 4 parejas (datos propios).
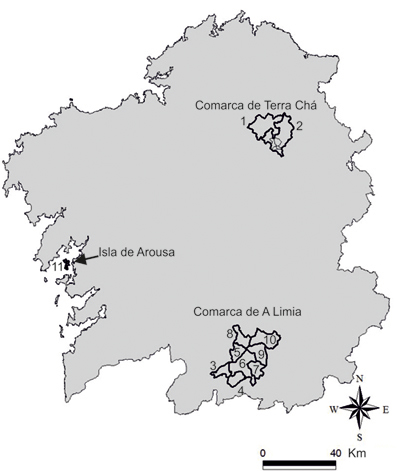
Tendencia poblacional A nivel europeo se considera en declive, con una tendencia de moderada disminución entre 1970 y 1990 identificada fundamentalmente en base a censos de invierno (Birdlife International, 2004a). A tenor de los datos disponibles, la tendencia de la población nidificante española se puede caracterizar por una contracción del área de distribución y la reducción de efectivos nidificantes. En Asturias fue reproductor habitual durante la década de 1970, desconociéndose si también lo fue con anterioridad. Nidificó en las cercanías de la ría de Villaviciosa (Noval, 1975), con 1-2 parejas en el monte Cubera desde 1971 a 1977 (Noval y Cortés, 2000), y también en la orilla asturiana de la ría de Ribadeo, donde se mencionó una única pareja. Desde la década de 1980 hasta la actualidad sólo se ha documentado una posible pareja reproductora en Santolaya (Gozón) (García, 2000). En las marismas del Guadalquivir se señalaron posibles intentos de cría anteriores a 1966 (Bernis, 1966; Glutz Von Blotzheimet al., 1986) y en diversas localidades del centro y sur de España se citó la presencia primaveral de ejemplares (Gullick y Jiménez, 1991; Santiago y Ortega, 1993; Hernández, 1997; Chiclana, 2001; Rodríguez, 2012). Estas observaciones probablemente correspondieron en su gran mayoría a migrantes tardíos sedimentados o individuos veraneantes no reproductores, con la excepción quizás de la presencia de efectivos en la laguna del camino de Villafranca (Alcázar de San Juan, Ciudad Real) en la primavera de 1991, donde se escucharon cantos de celo y se vieron dos ejemplares en abril y junio (Gullick y Jiménez, 1991). En Galicia ha existido una población nidificante regular desde, al menos, el primer tercio del siglo XX (Tait, 1924; Villarinoet al., 2002), con presencia de efectivos reproductores en la antigua laguna de Antela (Ourense) en el periodo 1930-1960 (Villarinoet al., 2002). En estas décadas anteriores a 1960 pudo también nidificar en la comarca de Terra Chá (Lugo). En este sentido, aunque no hay datos concretos de cría, la alusión a posibles intentos de reproducción esporádicos en el NW ibérico (Bernis, 1966) quizás se refería a esta comarca gallega, ya que el autor residió en Lugo en la década de 1940 y prospectó en esa época esta comarca (Bernis, 1956). Hasta la fecha se ha documentado la nidificación del Zarapito real en tres zonas del territorio gallego: isla de Arousa (Pontevedra), comarca de A Limia (Ourense) y comarca de Terra Chá (Lugo), si bien en la actualidad sólo persiste la cría en esta última (Figura 3).
Figura 3. Municipios con presencia primaveral o reproducción confirmada, pasada y actual, de Zarapito real en Galicia. Municipios con nidificación confirmada: 1. Cospeito; 2. Castro de Rei; 9. Sarreaus; 10. Vilar de Barrio; 11. Isla de Arousa. Municipios con presencia primaveral de ejemplares: 3. Porqueira; 4. Os Blancos; 5. Sandiás; 6. Xinzo de Limia; 7. Trasmiras; 8. Xunqueira de Ambia. (Domínguez y Vidal, 2009).
En la isla de Arousa es un invernante común, con frecuentes observaciones de ejemplares sedimentados en primavera (Domínguez, 1988) que no implican episodios de cría. El único evidenciado hasta la fecha se produjo en 1988, con una pareja reproductora localizada en el sur de la isla (Bárcena y Domínguez, 1995). En la comarca de A Limia la nidificación estuvo ligada a la antigua laguna de Antela y su entorno, laguna que fue desecada a finales de la década de 1950 (Martínez, 1997b; Villarinoet al., 2002). La presencia de ejemplares en periodo reproductivo y los episodios de nidificación se documentaron en 8 ayuntamientos de esta comarca: Porqueira, Rairiz de Veiga, Sandiás, Sarreaus, Trasmiras, Vilar de Barrio, Xinzo de Limia y Xunqueira de Ambia (Figura 3). En el periodo 1930-1960 esta limícola presentó una distribución local más amplia y con mayores efectivos reproductores que en décadas posteriores. En ese periodo se confirmó la nidificación en el ayuntamiento de Sarreaus (Villarinoet al., 2002), si bien las abundantes referencias de presencia de ejemplares en meses primaverales sugieren la posibilidad de eventos de cría en otros puntos de la periferia de la laguna (Villarino et al., 2002). Desde 1975 sólo se ha mencionado la reproducción en 1983, año en el que se censaron 3-5 parejas en el ayuntamiento de Vilar de Barrio (Domínguezet al., 1987; Villarinoet al., 2002). Posteriormente sólo se han realizado escasas observaciones primaverales, sin indicios de cría, en los ayuntamientos de Porqueira y Xunqueira de Ambia (Villarinoet al., 2002). En la comarca de Terra Chá parece probable que haya sido un reproductor habitual a lo largo de buena parte del siglo XX, existiendo entre 1960 y 1980 mención de posibles intentos de reproducción (Domínguezet al., 1987; Martínezet al., 2004). Los primeros datos concretos de cría datan de la década de 1980, localizándose en el entorno de la laguna de Cospeito (Domínguezet al., 1987; Castro y Castro, 1990). Esta laguna fue desecada a finales de la década de 1950 y restaurada, con morfología diferente, a principios de la década de 2000 (Ramil y Domínguez, 2006). Desde comienzos de la década de 1990 se han sucedido las observaciones de cría, todas ellas en los ayuntamientos de Cospeito y Castro de Rei (Martínez y Salaverri, 1994; Salaverri, 1994; Martínez, 1995a, 1995b, 1996, 1997a; Salaverri, 1999). Desde 2007 la población nidificante se ha ubicado en el ayuntamiento de Castro de Rei, aunque con presencia primaveral de ejemplares en otras zonas de este ayuntamiento y de los de Cospeito y Begonte. En resumen, la contracción espacial del área de distribución de la especie en España se evidenció a partir de la desecación de la laguna gallega de Antela (Ourense), acontecida en la segunda mitad de la década de 1950, y se agudizó con la desaparición del núcleo de cría asturiano hacia finales de la década de 1970. Respecto a la tendencia del número de parejas reproductoras en España, pueden definirse dos periodos desde el primer cuarto del siglo XX: uno anterior a 1960, caracterizado por la existencia de las lagunas gallegas de Antela y de Cospeito, ambas desecadas a finales de la década de 1950 (Martínez, 1997b; Ramil y Domínguez, 2006), y otro desde 1960 a la actualidad (Domínguez y Vidal, 2009).
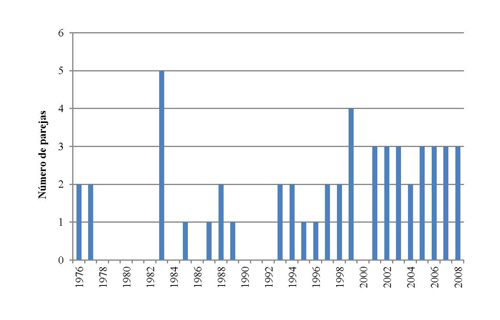
Figura 4. Número mínimo de parejas reproductoras de Zarapito real en Galicia desde mediados de la década de 1970. Los años sin censo reflejan ausencia de datos propios o publicados (Domínguez y Vidal, 2009). El valor asignado a 1983 corresponde al más elevado del rango estimado para ese año (ver texto).
Con anterioridad a 1960 la población nidificante pudo estar constituida por un mínimo de 10 parejas, aparentemente todas localizadas en Galicia (Domínguez y Vidal, 2009), hipótesis sustentada en la extensión de humedales apropiados en las comarcas gallegas de A Limia y Terra Chá y la información concreta publicada para la comarca de A Limia (Villarinoet al., 2002). El periodo comprendido desde 1960 a la actualidad se caracteriza por una total ausencia de datos en la década de 1960, siendo imposible determinar la posible distribución y abundancia en estos años, y la existencia de datos cuantitativos de calidad variable desde la década de 1970 (Domínguez y Vidal, 2009). A mediados de la década de 1990 se estimó una población en el N de España de 6-12 parejas (Bednorz y Grant, 1997). Entre 1975 y 2008 la población anual mínima conocida para Galicia osciló entre 1 y 5 parejas (Figura 4), si bien la población real probablemente fue mayor en algunos de los años de la serie ya que, por ejemplo, para 1983, en el que se observaron 3-5 parejas reproductoras en la Limia, no se dispuso de datos referidos a Terra Chá. A esta población gallega habría que añadir la reproductora en Asturias durante buena parte de la década de 1970, aparentemente no superior a 5 parejas/año. En todo caso, a tenor de la ubicación de la Península en el límite meridional de distribución de la especie, no parece probable que la población española haya sido numerosa a lo largo del siglo XX, pudiendo estimarse en un orden de magnitud de muy pocas decenas de parejas (Domínguez y Vidal, 2009).
Estado de conservación Categoría global IUCN (2012): Casi Amenazada NT (BirdLife International, 2013). Categoría IUCN para España (2004): En Peligro EN (Martínez et al., 2004). Especie incluida en el Anexo II/B de la Directiva 2009/147/CEE relativa a la conservación de las aves silvestres, en el Anexo III de la Convención de Berna y en el Anexo II de la Convención de Bonn, relativa a la Conservación de especies silvestres migratorias. La población nidificante gallega figura en el Catálogo Nacional de Especies Amenazadas en la categoría de “En peligro de extinción” (Real Decreto 139/2011). A nivel autonómico, la población nidificante gallega ha sido incluida en la categoría de “En peligro de extinción” en el Catálogo Gallego de Especies Amenazadas (Decreto 88/2007, Xunta de Galicia). En Asturias figura como “Vulnerable” en el Catálogo Regional de Especies Amenazadas de la Fauna Vertebrada del Principado de Asturias (Decreto 32/90). Esta Comunidad Autónoma también ha promulgado el Plan de Conservación del Zarapito Real en dicho Principado (Decreto 103/2002). Es considerada de “Interés especial” en los Catálogos Regionales de Especies Amenazadas de Extremadura (Decreto 37/2001) y de Castilla la Mancha (Decreto 33/1998). Ha sido clasificada como SPEC 2, categoría que engloba aquellas especies caracterizadas por estar presentes principalmente en Europa y con estado de conservación desfavorable en nuestro continente (Birdlife International, 2004a, 2004b).
Amenazas Sobre la población nidificante española se cierne un conjunto de amenazas de importancia variable, entre las que cabe mencionar: 1) Cambios en el uso del suelo (destrucción y alteración del hábitat). Relacionados fundamentalmente con la modificación, o manejo inadecuado, de áreas de matorral utilizadas por los efectivos reproductores y la transformación de pastizales higrófilos y praderías en cultivos de maíz (Zea mays). Algunos de los parches de matorral ubicados en Terra Chá donde nidificó el zarapito han sido repoblados con coníferas y eucaliptos. En este sentido, aunque se ha citado su reproducción en medios provistos de pinos de hasta 6-7 años (Dubois y Mahéo, 1986), el crecimiento de los árboles termina expulsando a las parejas nidificantes al hacer el hábitat inapropiado (Cramp y Simmons, 1993; Dubois y Mahéo, 1986). Además de la reforestación de zonas de cría, el desarrollo excesivo del tojo (Ulex europaeus) también constituye una amenaza, ya que la ausencia de manejo regular mediante cortas periódicas, con el consiguiente crecimiento del matorral, expulsa a los adultos de áreas potenciales de nidificación (Dubois y Mahéo, 1986). Esta disminución de la superficie de matorral en beneficio de las repoblaciones forestales se ha constatado en el conjunto de la comarca de Terra Chá, y particularmente en los ayuntamientos de Cospeito y Castro de Rei, entre 1956 y 2004 (Corbelle y Crecente, 2008). En el conjunto de la comarca de Terra Chá, entre 1941 y 1971 la superficie repoblada con árboles fue de unas 16.000 ha (Rico-Boquete, 1995). Otra amenaza de transformación, planteada en años recientes sobre el área de reproducción existente en Castro de Rei, aunque afortunadamente no ejecutada, ha sido la referida a la instalación de un centro de desguace de aviones. En el pasado se han transformado pastizales higrófilos, naturales o seminaturales, y praderías de Cospeito y Castro de Rei en cultivos de maíz, siendo estos desfavorables para la presencia de zarapitos. Cambios similares se produjeron también en las últimas dos décadas en zonas de la comarca de A Limia (Ourense) donde nidificó. 2) Perturbaciones de origen humano. Durante el periodo reproductivo se producen vuelos de aeronaves, fundamentalmente avionetas, en el aeródromo de Rozas, adyacente al área actual de cría. Los vuelos de aeronaves constituyen una fuente potencial de perturbaciones sobre limícolas y otras aves (De Roos, 1983; Barryet al., 1985; Kempf y Hüppop, 1995; Schuecket al., 2001), siendo especialmente impactantes los vuelos de pequeños aviones a baja altura (< 500 m) (Belanger y Bedard, 1989; Liddle, 1997). A pesar del impacto potencial descrito para diferentes especies, la información recogida en el periodo 2007-2012 sugiere una incidencia muy limitada o incluso inexistente de estos vuelos de avionetas sobre el comportamiento de los adultos incubantes y las familias con pollos. Sin embargo, otro tipo de aeronaves como helicópteros, de presencia hasta ahora esporádica en dicho aeródromo, y el uso del entorno para actividades de aeromodelismo podrían suponer una fuente potencial de perturbaciones de consecuencias difíciles de predecir. En localidades de reproducción alemanas se ha relacionado la existencia de perturbaciones por vuelos de aviones de aeromodelismo con la interrupción del proceso de incubación y reducción del área de campeo (Boschert, 2004). La actual zona de ubicación de nidos se caracteriza por presentar un tránsito de personas muy reducido (datos propios), ligado al desbroce ocasional de parches de matorral para uso ganadero. Con todo, el tránsito de gente constituye una fuente potencial de perturbaciones para limícolas reproductores, con posible incidencia en su éxito reproductivo (Liddle, 1997), por lo que la viabilidad de este núcleo puede depender críticamente de la ausencia de molestias inducidas por personas. El área de campeo de adultos y pollos en crecimiento se enmarca en un área con pistas asfaltadas y caminos con tránsito humano relacionado fundamentalmente con actividades agrícolas. Al igual que en el caso de la zona de ubicación de nidos, los niveles actuales de tránsito de vehículos por las pistas y de personas no parecen incidir de forma relevante en las aves adultas y pollos (datos propios), si bien serían necesarios estudio pormenorizados al respecto. 3) Actividades agrícolas. Pueden tener sobre las poblaciones de limícolas efectos directos, en términos de pérdida de nidos y pollos en crecimiento, e indirectos (Macdonald y Bolton, 2008). La evidencia disponible parece indicar que las tasas de mortalidad en nido por depredación pueden incrementarse con la intensificación agrícola, pero esta cuestión necesitaría de estudios adicionales (Macdonald y Bolton, 2008). Las zonas de presencia y reproducción del Zarapito real en Terra Chá se caracterizan por un predominio de granjas lácteas con ganado estabulado, en las que una parte sustancial del alimento del ganado se obtiene mediante la recolección de la hierba a lo largo de la primavera y verano. La fenología de recolección puede estar condicionada por la meteorología local, interaccionando así en distintos grados, según años, con la reproducción del zarapito. La utilización de segadoras mecánicas en prados donde se ubican nidos o se mueven pollos no volantones puede ocasionar mortalidad (Kruket al., 1996). En los últimos años, los nidos en Terra Chá se han encontrado en parches de landa no sometidos a pastoreo, pero hay constancia de desaparición de nidos ubicados en prados (Martínez, 1995b) y de pollos no volantones (datos propios) como consecuencia de la siega mecánica de prados. Por el contrario, a diferencia de lo señalado en otras poblaciones europeas de limícolas de pastizal (Beintema y Muskens, 1987; Durant et al., 2008), la mortalidad en nido por pisoteo de ganado ha sido inexistente en los últimos años en Terra Chá, ya que la zona de ubicación de nidos no ha sido sometida a pastoreo. 4) Depredación. La abundancia y comportamiento de los depredadores juega un papel muy importante en la mortalidad en nido de limícolas (Macdonald y Bolton, 2008). La depredación de nidos y pollos no volantones constituye una de las principales causas de mortalidad en poblaciones europeas de Zarapito real (Grantet al., 1999; Valkamaet al., 1999; Englet al., 2004; Boschert, 2005; Grimm, 2005). Los depredadores de nidos más frecuentes son mamíferos, sobre todo zorros (Vulpes vulpes, Grantet al., 1999; Brandsma, 2002; Englet al., 2004; Boschert, 2005). Las aves, fundamentalmente córvidos, tienen una incidencia desigual según localidades, ya que en algunas se ha evidenciado como depredadoras importantes de nidos (Grantet al., 1999), mientras que en otras su efecto ha sido prácticamente inexistente (Boschert, 2005). Los datos disponibles para la población española revelan una importante mortalidad en nido (36,9 % de nidos depredados en el periodo 2007-2012), con especial incidencia de depredadores de actividad nocturna, muy probablemente mamíferos (datos propios). La zona de cría se caracteriza también por la presencia primaveral de varias Falconiformes, destacando como área de reproducción de 2-3 parejas de Aguilucho cenizo (Circus pygargus). 5) Incendios. Las áreas de matorral son susceptibles de sufrir incendios, con consecuencias potencialmente negativas para la reproducción (Martínezet al., 2004). En los últimos años en la zona de cría se produjeron incendios ocasionales en primavera y verano. No se constató su afección en nidos activos o pollos, pero sí ardieron parches donde en temporadas precedentes se ubicaron nidos (datos propios). 6) Contaminación. La población nidificante gallega se ubica en un medio agrícola caracterizado por el uso frecuente de fertilizantes y productos fitosanitarios. En otras zonas europeas se han detectado elevadas concentraciones de contaminantes en huevos de Zarapito real, tanto de compuestos organoclorados (Boschert, 1992; Meiseret al., 2003; Boschert, 2004) como de metales pesados (Currie y Valkama, 1998). Con relación a las concentraciones de metales pesados en huevos (Cu, Ni, Zn, Pb, Cd), en un estudio efectuado en áreas agrícolas de Finlandia aparentemente no hubo diferencias entre áreas contaminadas y no contaminadas ni en el peso de adultos incubantes (Currie y Valkama, 1998) ni en las concentraciones de calcio, pero las cáscaras de huevos de áreas contaminadas tuvieron mayores concentraciones de metales pesados (Currie y Valkama, 1998). La conclusión de este estudio fue que la contaminación por metales pesados tuvo efectos inmediatos mínimos en la alimentación de adultos y en su éxito reproductivo (Currie y Valkama, 1998). Sin embargo, otros estudios han evidenciado efectos indirectos de pesticidas y herbicidas, tanto en los invertebrados presa como en el propio éxito reproductivo de especies ligadas a medios agrícolas (Boatmanet al., 2004). Otra fuente potencial de contaminación puede provenir de incendios forestales generadores de hidrocarburos policíclicos aromáticos (PAH) de origen pirogénico, los cuales pueden ser incorporados a los huevos (Vidalet al., 2011). En el área de cría de Terra Chá no hay datos sobre niveles de contaminantes en el suelo, ni en las especies presa ni en huevos de zarapito. 7) Pequeño tamaño poblacional. Las poblaciones amenazadas, en virtud de su pequeño tamaño y aislamiento geográfico, presentan un riesgo de extinción considerablemente mayor que poblaciones numerosas (Caughley y Gunn, 1996). Otros factores de riesgo asociados a un reducido tamaño poblacional son la estocasticidad demográfica, cambios azarosos en el sex-ratio y dificultad para formar pareja (efecto Allee, USFWS, 1996). En este sentido, la elevada filopatría del Zarapito real y el aislamiento geográfico de esta población española incidirían en la posible magnificación de estos riesgos.
Medidas de conservación La especie está estrictamente protegida en España. El área actual de cría se encuentra dentro de la Reserva de la Biosfera “Terras do Miño”, declarada en noviembre de 2002. Hasta la fecha no se han adoptado medidas de manejo y conservación del hábitat de nidificación o de las zonas de alimentación de adultos reproductores y pollos en crecimiento. Martínez et al. (2004) han propuesto medidas de protección y vigilancia, investigación aplicada y seguimiento de la población reproductora. El mantenimiento de prácticas tradicionales de recolección de invertebrados durante la marea baja es compatible con la presencia de poblaciones invernantes de Zarapito real. Un estudio realizado en el Parque Natural de las Marismas de Santoña en el que se compararon periodos anteriores y posteriores a la recolección no encontró diferencias significativas en la abundancia de Zarapitos reales, su actividad de búsqueda de alimento, el porcentaje de cangrejos en la dieta y la talla de los cangrejos (Navedo y Masero, 2008). Se ha recomendado reducir el número de recolectores durante el otoño a 0,56 personas/10 ha-1 (Navedo y Masero, 2007).
Referencias Arévalo Baca, J. (1887). Aves de España. Viuda e Hijo de Aguado, Madrid. Atkinson, P. W., Fuller, R. J., Vickery, J. A. (2002). Large-scale patterns of summer and winter bird distribution in relation to farmland type in England and Wales. Ecography, 25: 466-480. Bárcena, F., Domínguez, J. (1995). Numenius arquata. Pp. 418. En: S.G.H.N. (Ed.). Atlas de Vertebrados de Galicia. Tomo II. Aves. Consello da Cultura Galega, Santiago de Compostela. Barry, T., Everitt, R., Johnson, S. (1985). Beaufort Environmental Monitoring Project 1983-1984. Hypothesis nº 12. The effects of low altitude aircraft flighs on staging brant. Environ. Stud. (Ott.), 34: 175-177. Bednorz, J., Grant, M. (1997). Numenius arquata. Pp. 300-301. En: Hagemeijer, W. J. M., Blair, M. J. (Eds.). The EBCC Atlas of European Breeding Birds. T & AD Poyser, London. Beintema, A. J., Muskens, G. J. D. M. (1987). Nesting success of birds breeding in Dutch agricultural grasslands. Journal of Applied Ecology, 24 (3): 743-758. Belanger, L., Bedard, J. (1989). Responses of staging greater snow geese to human disturbance. Journal of Wildlife Management, 53 (3): 713-719. Bernis, F. (1954). Prontuario de la avifauna Española. Ardeola, 1: 11-85. Bernis, F. (1956). Nota preliminar sobre aves de Asturias y Galicia. Ardeola, 3 (1): 31-49. Bernis, F. (1966). Aves Migradoras Ibéricas. Fascículo 4º. Sociedad Española de Ornitología. Madrid. Birdlife International (2004a). Birds in Europe: population estimates, trends and conservation status. Birdlife International. Wageningen. Birdlife International (2004b). Birds in the European Union: a status assessment. Birdlife International. Wageningen. BirdLife International (2013). Numenius arquata. En: IUCN Red List of Threatened Species. Version 2013.1. <www.iucnredlist.org>. Boatman, N. D., Brickle, N. W., Hart, J. D., Milsom, T. P., Morris, A. J., Murray, A. W. A., Murray, K. A., Robertson, P. A. (2004). Evidence for the indirect effects of pesticides on farmland birds. Ibis, 146 (Suppl. 2): 131-143. Boschert, M. (1992). Ruckstande chlororganischer Verbindungen in Gelegen des Grossen Brachvogels (Numenius arquata) aus einem Brutgebiet am sudlichen Oberrhein. Ökol. Vögel., 14: 101-111. Boschert, M. (2004). Der Gross e Brachvogel (Numenius arquata [Linnaeus 1758]) am badischen Oberrhein - Wissenschaftliche grundlagen fur einen umfassenden und nachhaltigen schutz. University of Tubingen. Boschert, M. (2005). Gelegeverluste beim grossen brachvogel Numenius arquata am badischen oberrhein - ein vergleich von 2000-2002 mit fruheren zeitraumen unter besonderer berucksichtigung der pradation. Vogelwelt, 126 (4): 321-332. Brandsma, O. (2002). The influence of the Fox on meadow birds in a wetland in NW-Overijssel. Levende Natuur, 103 (4): 126-131. Broyer, J., Roche, J. (1991). La population nicheuse de Courlis Cendré Numenius arquata du Bassin de la Saône. Alauda, 59 (3): 129-135. Butiev, V. T., Lebedeva, E. A. (1998). Curlew Numenius arquata in the Vologda region of north-European Russia. Pp. 299-302. En: H. Hötker, E. Lebedeva, P. S. Tomkovich, J. Gromadzka, N. C. Davidson, Evans, L., Stroud, D. A., West, R. B. (Eds.). Migration and international conservation of waders. Research and conservation on North Asian, African and European flyways. International Wader Studies 10. Castro, J., Castro, X. C. (1990). As Lagoas de Cospeito. Introducción ó estudio dun humidal. Servicio de Publicaciones. Diputación Provincial de Lugo, Lugo. Catry, P., Costa, H., Elias, G., Matías, R. (2010). Aves de Portugal. Ornitología do território continental. Assirio & Alvim, Lisboa. Caughley, G., Gunn, A. (1996). Conservation biology in theory and practice. Blackwell Scientific, Cambridge. Corbelle, E., Crecente, R. (2008). Estudio da evolución da superficie agrícola na comarca da Terra Chá a partir de fotografía aérea histórica e mapas de usos, 1956-2004. Recursos Rurais, 1 (4): 57-65. Cramp, S., Simmons, K. E. L. (Eds.) (1993). Handbook of the Birds of Europe the Middle East and North Africa. The Birds of the Western Palearctic. Volume III. Waders to Gulls. Oxford University Press, Oxford. Currie, D., Valkama, J. (1998). Limited effects of heavy metal pollution on foraging and breeding success in the curlew (Numenius arquata). Environmental Pollution, 101: 253-261. Currie, D., Valkama, S. K. (2000). Population density and the intensity of paternity assurance behaviour in a monogamous wader: the Curlew Numenius arquata. Ibis, 142 (3): 372-381. Chiclana, F. (2001). Zarapito real Numenius arquata. Ardeola, 48 (1): 143. De Juana, E. (1997). Zarapito real Numenius arquata. Pp. 557. En: Purroy, F. J. (Ed.). Atlas de las Aves de España (1975-1995). Lynx, Barcelona. De Roos, G. T. (1983). Military training in the Wadden Sea area. Pp. 106-112. En: Bruyns, M. F. M., Wolff, W. J. (Eds.). Nature conservation, nature management and physical planning in the Wadden Sea area. Balkema, Rotterdam. Deceuninck, B. (2001). Breeding waders in France: populations, trends and distributions: 1984-1996. WSG Bulletin, 95: 45-50. Domínguez, J. (1988). Taxocenosis de limícolas de las rias gallegas. Con especial referencia a las de Arosa y Ortigueira. Tesis Doctoral. Santiago de Compostela. Domínguez, J. (1997). Invernada y migración de limícolas en el litoral atlántico ibérico. Pp. 35-75. En: Barbosa, A. (Ed.). Las aves limícolas en España. Organismo Autónomo Parques Nacionales, Madrid. Domínguez, J., Bárcena, F., Souza, J. A., Villarino, A. (1987). Breeding waders in Galicia, north-west Spain. WSG Bulletin, 50: 28-29. Domínguez, J., Vidal, M. (2009). Zarapito real. Pp. 130-141. En: Palomino, D., Molina, B. (Eds.). Aves acuáticas reproductoras. Población en 2007 y método de censo. SEO/BirdLife, Madrid. Dubois, P. J., Mahéo, R. (1986). Limicoles Nicheurs de France. Ministere de l'Environnement, Paris. Durant, D., Tichit, M., Kernêis, E., Fritz, H. (2008). Management of agricultural wet grasslands for breeding waders: integrating ecological and livestock system perspectives-a review. Biodiversity and Conservation, 17: 2275-2295. Engl, M., Leibl, F., Mooser, K. (2004). Bestandsentwicklung, Brutbiologie und Reproductionserfolg des Grossen Brachvogels Numenius arquata im Mettenbacher und Griessenbacher Moos, Landkreis Landshut. Ornithologischer Anzeiger, 43 (3): 217-235. García, E. (2000). Numenius arquata. Pp. 64. En: Álvarez-Balbuena, F., Vigil, A., Álvarez, C. M., Carballal, M. E., García, E., García, J. A. (Eds.). Aves raras y escasas en Asturies. Coordinadora Ornitolóxica d'Asturies, Gijón. Glutz von Blotzheim, U. N., Bauer, K. M., Bezzel, E. (1986). Handbuch der Vögel Mitteleuropas. Band 7. Charadriiformes (2. Teil). 2., durchgesehene Auflage. Aula Verlag, Wiesbaden. González, R., Pérez-Aranda, D. (2011). Las aves acuáticas en España, 1980-2009. SEO/BirdLife, Madrid. Grant, M. C., Orsman, C., Easton, J., Lodge, C., Smith, M., Thompson, G., Rodwell, S., Moore, N. (1999). Breeding success and causes of breeding failure of curlew Numenius arquata in Northern Ireland. Journal of Applied Ecology, 36 (1): 59-74. Grimm, M. (2005). Bestandsentwicklung und Gefaehrdungsursachen des Grossen Brachvogels Numenius arquata in den Belziger Landschaftswiesen (Brandenburg). Vogelwelt, 126 (4): 333-340. Gullick, T. M., Jiménez, J. (1991). Zarapito real Numenius arquata. Ardeola, 38 (2): 331. Hernández, A. J. (1997). Zarapito real Numenius arquata. Ardeola, 44 (2): 253. Iglesias, L. (1927). As aves de Galicia. Arquivos do Seminario de Estudos galegos, 1: 251-262. Issa, N., Boutin, J.-M. (2010). Anatidés et Limicoles nicheurs en France: enquêtes 2010. Présentation et méthodologie. Ministère de l’Ecologie, de l’Energie, du Développement Durable et de la Mer, Paris. Jubete, F. (Ed.) (2005). Anuario Ornitológico de Palencia. Volumen 0. (1998-2001). Asociación de Naturalistas Palentinos, Palencia. Kempf, N., Hüppop, O. (1995). Behaviour of meadow birds towards aircraft close to an airport. WSG Bulletin, 76: 21. Kruk, M., Noordervliet, M. a. W., Ter Keurs, W. J. (1996). Hatching dates of waders and mowing dates in intensively exploited grassland areas in different years. Biological Conservation, 77: 213-218. Lebedeva, E. A. (1998). Waders in agricultural habitats of European Russia. Pp. 315-324. En: Hötker, H., Lebedeva, E., Tomkovich, P. S., Gromadzka, J., Davidson, N. C., Evans, L., Stroud, D. A., West, R. B. (Eds.). Migration and international conservation of waders. Research and Conservation on North Asian, African and European flyways. International Wader Studies 10. Liddle, M. J. (1997). Recreation ecology. Chapman & Hall, London. López, Z., Guitián, J. (1983). Atlas provisional de los vertebrados terrestres de Galicia. Aves nidificantes. Universidad de Santiago, Santiago de Compostela. López Seoane, V. (1886). Reseña de la Historia Natural de Galicia. Imprenta Soto Freire, Lugo. Macdonald, M. A., Bolton, M. (2008). Predation on wader nests in Europe. Ibis, 150: 54-73. Madroño, A., González, C., Atienza, J. C. (2004). Libro Rojo de las Aves de España. Dirección General para la Biodiversidad-SEO/Birdlife, Madrid. Martínez, G. (1995a). Numenius arquata, Mazarico curlí. Pp. 31. En: Salaverri, L. J., Munilla, I. (Eds.). Segundo Anuario das Aves de Galicia. Sociedade Galega de Ornitoloxía, Santiago de Compostela. Martínez, G. (1995b). Zarapito real Numenius arquata. Ardeola, 42 (2): 222. Martínez, G. (1996). Numenius arquata, Mazarico curlí. Pp. 43. En: Salvadores, R., Vidal, C. (Eds.). III Anuario das Aves de Galicia. Sociedade Galega de Ornitoloxía, Santiago de Compostela. Martínez, G. (1997a). Zarapito real Numenius arquata. Ardeola, 44 (2): 253. Martínez, G., Salaverri, L. J. (1994). Zarapito real Numenius arquata. Ardeola, 41 (1): 98. Martínez, G., Vázquez, X., Mouriño, J., Salaverri, L. J. (2004). Zarapito Real Numenius arquata. Pp. 236-237. En: Madroño, A., González, C., Atienza, J. C. (Eds.). Libro Rojo de las Aves de España. Dirección General para la Biodiversidad-SEO/Birdlife, Madrid. Martínez, X. L. (Ed.) (1997b). Antela. A memoria asolagada. Xerais, Vigo. Meiser, H., Hagedorn, H. W., Ruf, J., Schulz, R. (2003). Antiandrogen p,p '-DDE, principal metabolite of DDT and PCBs in curlew eggs (Numenius arquata). Tierarztliche Umschau, 58 (3): 155-156. Mischenko, A. L., Sukhanova, O. V. (1998). Waders of the Novgorod region: peculiarities of their distribution and important breeding areas. Pp. 285-290. En: Hötker, H., Lebedeva, E., Tomkovich, P. S., Gromadzka, J., Davidson, N. C., Evans, L., Stroud, D. A., West, R. B. (Eds.). Migration and international conservation of waders. Resarch nad conservation on North Asian, African and European flyways. International Wader Studies 10. Navedo, J. G., Arranz, D., Herrera, A. G.; Salmon, P., Juanes, J. A., Masero, J. A. (2013). Agroecosystems and conservation of migratory waterbirds: importance of coastal pastures and factors influencing their use by wintering shorebirds. Biodiversity and Conservation, 22 (9): 1895-1907. Navedo, J., Masero, J. A. (2007). Measuring potential negative effects of traditional harvesting practices on waterbirds: a case study with migrating curlews. Animal Conservation, 10 (1): 88-94. Navedo, J., Masero, J. A. (2008). Effects of traditional clam harvesting on the foraging ecology of migrating curlews (Numenius arquata). Journal of Experimental Marine Biology and Ecology, 355 (1): 59-65. Noval, A. (1975). El libro de la Fauna ibérica. Aves. Naranco, Oviedo. Noval, A., Cortés, X. (2000). Numenius arquata. Pp. 64. En: Álvarez-Balbuena, F., Vigil, A., Álvarez, C. M., Carballal, M. E., García, E. (Eds.). Aves raras y escasas en Asturias. Coordinadora Ornitolóxica d'Asturies, Gijón. O'Brien, M., Bainbridge, I. (2002). The evaluation of key sites for breeding waders in lowland Scotland. Biological Conservation, 103: 51-63. Ramil, P., Domínguez, J. (Eds.) (2006). A lagoa de Cospeito. Consellería de Medio Ambiente e Desenvolvemento Sostible, Santiago de Compostela. Reed, T. M., Langslow, D. R., Symonds, F. L. (1983). Breeding waders of the Caithness flows. Scottish Birds, 12 (6): 180-186. Reyes y Prosper, V. (1886). Catálogo de las aves de España, Portugal e Islas Baleares. Anales de la Real Sociedad Española de Historia Natural, 15 (1): 5-109. Rico-Boquete, E. (1995). Política forestal e repoboacións en Galicia (1941-1971). Servicio de Publicacións e Intercambio Científico, Universidade de Santiago de Compostela, Santiago de Compostela. Ríos Naceyro, F. (1850). Catálogo de las aves observadas en las cercanías de Santiago y otros puntos de Galicia. Memoria de la Real Academia de las Ciencias, 1: 93-116. Rodríguez, R. (2012). Chorlitejo grande y zarapito real en Palencia. Quercus, 314: 49. Salaverri, L. J. (1994). Numenius arquata, Mazarico curlí. Pp. 25. En: Munilla, I., Guitián, J. (Eds.). Primeiro Anuario das Aves de Galicia. Sociedade Galega de Ornitoloxía, Santiago de Compostela. Salaverri, L. J. (1999). Zarapito real Numenius arquata. Ardeola, 46 (1): 157. Santiago, J. M., Ortega, F. (1993). Zarapito real Numenius arquata. Ardeola, 40 (1): 96. Scott, D. (2009). Eurasian Curlew Numenius arquata. Pp. 307-311. En: Delany, S., Scott, D. A., Dodman, T., Stroud. D. A. (Eds.). An Atlas of Wader Populations in Africa and Western Eurasia. Wetlands International, Wageningen. Schueck, L. S., Marzluff, J. M., Steenhof, K. (2001). Influence of military activities on raptor abundance and behavior. Condor, 103 (3): 606-615. Tait, W. C. (1887). A List of Birds of Portugal. Ibis, 29: 372-400. Tait, W. C. (1924). The Birds of Portugal. Witherby, London. Thorup, O. (2006). Breeding waders in Europe 2000. International Wader Studies 14. Tuule, E., Tuule, A., Elts, J. (2003). Suurkoovitaja pesitsusaegsest arvukusest saue seirealal aastatel 1963-2002. Hirundo, 16 (1): 14-22. USFWS (1996). Piping Plover (Charadrius melodus), Atlantic Coast population, revised recovery plan. Hadey, Massachusetts. Valkama, J., Currie, D. (1999). Low productivity of Curlews Numenius arquata on farmland in southern Finland: Causes and consequences. Ornis Fennica, 76 (2): 65-70. Valkama, J., Currie, D., Korpimaki, I. (1999). Differences in the intensity of nest predation in the curlew Numenius arquata: A consequence of land use and predator densities? Ecoscience, 6 (4): 497-504. Vidal, M., Domínguez, J., Luis, A. (2011). Spatial and temporal patterns of polycyclic aromatic hydrocarbons (PAHs) in eggs of a coastal bird from northwestern Iberia after a major oil spill. Science of the Total Environment, 409: 2668-2673. Villarino, A., González, S., Bárcena, F. (2002). Vertebrados da Limia. I. Aves: Gaviiformes a Piciformes. Limaia, Sandiás. Wilson, A. M., Ausden, M., Milson, T. P. (2004). Changes in breeding wader populations on lowland wet grasslands in England and Wales: causes and potential solutions. Ibis, 146 (Suppl. 2): 32-40. Wylegala, P., Wylegala, S., Pinkowski, R., Kujawa, D. (2004). A decline in the Curlew Numenius arquata numbers in the Notec River valley in 1980-2004. Notatki Ornitologiczne, 45 (2): 120-122.
Jesús Domínguez y María Vidal Fecha de publicación: 28-10-2013 Domínguez, J., Vidal, M. (2016). Zarapito real - Numenius arquata. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Morales, M. B. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||