|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: Aurelio’s Rock Lizard, identification, description, measurements, variation.
Sinónimos y combinaciones Lacerta (Archaeolacerta) aurelioi Arribas, 1994. Archaeolacerta aurelioi – Arribas, 1997.
Origen y evolución La datación y secuencia de diferenciaciones entre las distintas especies de lagartijas pirenaicas, difieren según la aproximación genetica empleada. A partir del estudio de electroforesis de aloenzimas, Iberolacerta aranica comienza a diferenciarse tempranamente del antepasado común de I. aurelioi y I. bonnali, hace aproximadamente entre 2.25 y 1.55 (media de 2) millones de años respectivamente (Dnei= 0.45 respecto a I. bonnali y Dnei=0.31 respecto a I. aurelioi) coincidiendo con las fases iniciales del Pleistoceno mientras que estas últimas dos especies se habrían diferenciado hace 450.000 años (Dnei=0.09 entre I. aurelioi e I. bonnali) durante el Pleistoceno Medio. Las diferencias genéticas encontradas en la secuenciación del gen mitocondrial 12S rRNA son aproximadamente de un 2% entre las especies pirenaicas. Eso quiere decir que su diferenciación se remonta según esta técnica y este gen a unos 2 millones de años, es decir, al inicio de las glaciaciones pleistocénicas. Según el método de agregación (distancia o parsimonia -mínimo número de cambios posibles) la especie más diferenciada varía. Con el primero, por distancias, aparentemente es la lagartija pirenaica la más diferenciada, pero esto no está tan claro observando las distancias recíprocas entre ellas. Concretamente, la diferencia menor es entre la lagartija pirenaica (I. bonnali) y la pallaresa (I. aurelioi), que sería de 1.8 % (aprox. 1.8 millones de años de divergencia) y que casa bien con la hipótesis previas basadas en osteología y cariotipos. La distancia genética de la lagartija pallaresa a la aranesa, sería de 2.2% (2.2 millones de años de separación entre las poblaciones de Barlonguere y Orlà -pertenecientes a I. aranica- respecto a las de Mont-Roig -ya perteneciente a I. aurelioi-). La diferencia entre la aranesa (I. aranica) y la pirenaica (I. bonnali) es de 2.4% (2.4 millones de años). Si se toma como criterio el arbol de mínima extensión, también aparece la lagartija aranesa como la más diferenciada. Otro estudio que tiene en cuenta el gen mitocondrial Cyt b (citocromo b) arroja estas diferencias: 6.2 % entre la pirenaica y la pallaresa, 7.4 % entre la pallaresa y la aranesa, y 9.8 % entre la pirenaica y la aranesa. Si se toma en conjunto los tres genes (Cyt b+12s rRNA+CMos; este último, un gen nuclear de evolución muy lenta), las diferencias conservan la misma escala recíproca: 2.5 % entre la pirenaica y la pallaresa, 3 % entre la pallaresa y la aranesa, y 3.7% entre la pirenaica y la aranesa. Está claro que la barrera más importante es el curso del Garona, ya que las diferencias más importantes se acumulan en todos los casos entre la lagartija pirenaica (I. bonnali) hoy presente al S y W del Valle de Arán, en la Maladeta y Aigües-Tortes, y la lagartija aranesa (I. aranica) del Mauberme y sus contrafuertes. Esto debió suceder entre 2.4 millones de años (con el gen 12s rRNA) o alrededor de 4.2±1.4 millones de años según la combinación de todos los genes estudiados (3.7% de divergencia). Ambas fechas pueden correlacionarse con el inicio de las glaciaciones, es decir con la presencia intermitente de glaciar en el valle de la Garona, o con el modelado y entalladura fluvial de la cuenca de la Garona que es de finales del Terciario en la forma que lo conocemos hoy en día. La segunda barrera más importante es el estrecho margen "en filo de cuchillo" que separa el Tuc de Mill del Macizo de Mont-Roig (al menos, es la zona en la que actualmente no vive ninguna especie de este grupo de lagartijas), que separó a la lagartija aranesa de la pallaresa, cuyo aislamiento entre poblaciones debió darse entre 2.2 millones de años (gen 12s rRNA) y 3.8±1.4 millones de años con el combinado de genes (3% de divergencia). El aislamiento debe estar relacionado, al igual que en el caso anterior con el modelado del relieve (haciendose la zona alpina de crestas muy estrecha) probablemente por acción de los glaciares de la alta Noguera Pallaresa. La tercera barrera es el valle de la Noguera Pallaresa, también glaciar, con una situación y génesis muy parecido al del Garona, que separó las poblaciones de lagartija pirenaica de las de la pallaresa hace unos 1.8 millones de años (gen 12s rRNA) o bien cerca de 4.2±1.4 millones de años (un 2.5% de divergencia). En todos los casos está claro que el margen de diferencias entre ellas es muy pequeño, quedaron aisladas "casi a la vez" o sea, en un margen muy estrecho de tiempo, y en general parece que de forma muy ligada a la dinámica de aparición glaciar o de sobreexcavación por éstos de las altas cuencas de la Garona y la Noguera Pallaresa. El tratamiento de las distancias mutuas entre las especies pirenaicas no resuelve las relaciones de parentesco entre ellas de forma inequívoca. Según el algoritmo de agregación (el método de construcción de árbol), la lagartija aranesa aparece como especie hermana de las otras dos (la pallaresa y la pirenaica serían más parientas entre sí) o bien sólo de la lagartija pallaresa (siendo entonces la pirenaica la más diferenciada). La primera hipótesis: (aranesa (pirenaica+pallaresa)) coincide con los resultados más plausibles de la osteología y cromosomas, mientras que la segunda (pirenaica (pallaresa+aranesa)) coincide con la del estudio de los microsatélites del DNA (Mayer y Arribas, 1996; Arribas, 1997; Mayer y Arribas, 2003; Carranza et al., 2004; Crochet et al., 2004).
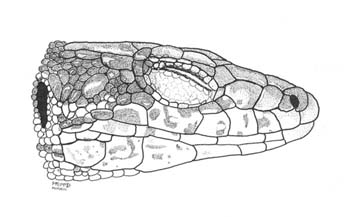
Descripción Folidosis Gránulos supraciliares en número de 1 a 14 (media = 6,5) en el lado derecho y 0 a 13 (6,4) en el lado izquierdo. Entre 20 y 30 (24,7) gularia; 7 a 15 (10,8) collaria; 37 a 52 (46,2) dorsalia; 25 a 29 (27,1) escamas ventrales en machos y 25 a 32 (29,2) en hembras; 7 a 16 (11,9) poros femorales en el lado derecho y 9 a 16 (12,1) en el derecho; 21 a 29 (24,9) lamellae bajo el cuarto dedo del pié; 6 a 10 (7,6) escamas circumanales. Escudos rostral e internasal generalmente en contacto (90,49 % de los ejemplares; escudos postocular y parietal generalmente separados (89,21 % de los ejemplares); escudos supranasal y loreal generalmente en contacto (92,91 % de los ejemplares). Hay una proporción elevada de anomalías (generalmente fragmentación) en las escamas cefálicas (Fig. 1).
Figura 1. Aspecto lateral de la cabeza. © O. Arribas.
Coloración Dorso de color de fondo pardo muy claro, a veces de tonos oliváceos en ejemplares que han mudado recientemente. Sobre este fondo aparecen dos bandas costales (= temporales) oscuras en los costados, y numerosas manchas oscuras de tamaño moderado a pequeño que en los machos y algunas hembras viejas llegan a recubrir todo el tracto dorsal y se unen a las bandas costales. Estas bandas se prolongan normalmente al menos en el tercio proximal de la cola. Diseño oscuro ventral muy marcado en el borde de las escamas, limitado a la parte más proximal y formando bandas transversales, más desarrollado hacia el vientre y región anal, generalmente sobre fondo amarillo yema. El patrón básico que corresponde a subadultos y hembras jóvenes presenta sólo las bandas costales sobre el fondo de color marrón muy claro, y se dan las siguientes variaciones ligadas al sexo y a la edad. Los recién nacidos (primer año calendario) tienen el dorso uniforme pero no es infrecuente un patrón oscuro de manchas. Partes inferiores blancas o algo amarillentas. Cola de color verde brillante. La coloración anarilla del vientre se hace visible al final de su primer año (segundo año calendario). Los subadultos (tercer año calendario) pueden tener o no la cola verdosa (aunque menos brillante que en los recién nacidos). El resto de su coloración es similar al patrón mencionado antes. Adultos (a partir del cuarto año calendario) con dimorfismo sexual en la coloración. Los machos tienen generalmente un abundante punteado oscuro en el dorso y píleo, que normalmente se prolonga por la parte superior de la cola. La banda lateral oscura inferior generalmente discontinua, formada por manchas. La coloración amarilla y las manchas oscuras del vientre están más desarrolladas en machos que en hembras. Los ejemplares viejos, tanto machos como hembras, están muy pigmentados. Las hembras viejas muestran más superficie amarilla, a veces algo anaranjada, que los machos viejos. Parte inferior de la cabeza y cuello de color blanco. Vientre y parte inferior de las patas de color amarillo. Parte inferior de la cola amarilla en hembras solamente (tanto más cuanto más viejas). En general, casi todos los adultos tienen el vientre con manchas oscuras. Solamente un 4,54 % de los ejemplares carece practicamente de manchas. El 8,67% tienen manchas bien desarrolladas solamente en las dos hileras más externas, el 18,59 % en las cuatro hileras más externas, y el 68,18 % en todas las hileras de ventrales. Los adultos, (especialmente los machos) tienen líneas claras dorsolaterales estrechas, no muy diferenciadas y sobre todo en la mitad anterior del dorso (Fig. 2 y 3) (Arribas, 1999a; 20141).
Figura 2. Hembra de lagartija pallaresa. © O. Arribas.
Figura 3. Aspecto ventral de la lagartija pallaresa. © O. Arribas.
Dimorfismo sexual Las proporciones de la cabeza y la longitud de miembros anteriores y posteriores son mayores en los machos (Arribas, 20141).
Tamaño Las hembras tienen mayor talla que los machos. La longitud media de cabeza y cuerpo de los machos es 52, 05 mm y la de las hembras 54,32 mm. La talla máxima de longitud de cabeza y cuerpo alcanza 65,25 mm en machos y 62,21 mm en hembras (Arribas, 1999a).
Cariotipo Formado por 26 cromosomas (10 metacéntricos y 16 acrocéntricos) en los machos y 25 (11 metacéntricos o submetacéntricos y 14 acrocéntricos) en las hembras. Sistema de cromosomas sexuales Z1Z2W. Bandas centroméricas bien marcadas en los cromosomas acrocéntricos y poco marcadas en los metacéntricos o submetacéntricos. NOR en posición telomérica en el brazo largo del tercer par de cromosomas (Odierna et al., 1996; Arribas, 1996 b).
Osteología Cráneo con ausencia por reducción secundaria de los procesos anterodistal del postfrontal y anteromedial del postorbitario. Postorbitario más largo que el postfrontal. Premaxilar con siete dientes y un processus nasalis esbelto y subparalelo. Maxilar con 15 a 17 dientes (media = 15,9) y dentario con 16 a 22 (media = 19,6). La mayoría de los dientes son bicúspides más que monocúspides. Los machos tienen generalmente 26 vértebras presacras y las hembras 27. Fórmula esternal-xiphisternal de tipo 3+2. Fontanela esternal ovalada, reducida secundariamente muy a menudo, ocasionalmente con uno o dos forámenes muy pequeños. Clavículas generalmente cerradas (emarginadas) e interclavícula típicamente cruciforme. Generalmente seis costillas dorsales posteriores cortas. Primeras vértebras caudales preautotómicas con procesos perpendiculares solamente en su parte anterior (Arribas, 1994, 1996a).
Hemipenes El aspecto general de los hemipenes es muy parecido a las de las otras especies de Iberolacerta. I. aurelioi tiene a igualdad de tamaño del animal, hemipenes sensíblemente más pequeños (y por tanto base de la cola menos ancha en los machos). Esto ha sido atribuido, a modo de hipótesis, a un posible caso de desplazamiento divergente de carácter por un contacto pasado con I. aranica, de cara a evitar la hibridación entre ambas. La microornamentación encontrada en esta especie es de tipo coroniforme (Arribas, 1994, 1996 b, 2007).
Datos aloencimáticos Esta especie difiere de I. aranica en tres alelos (AK, MDH-1 y GOT-1), y de I. bonnali en uno (PGM-2) (Mayer y Arribas, 1996).
Variación geográfica Se han observado pequeñas diferencias morfológicas y de folidosis entre las poblaciones de I. aurelioi, que parecen ser el resultado de un aislamiento reciente (Holoceno) a partir de un área más continua originada desde un unico refugio durante el Würm (Arribas, 1999a).
Referencias Arribas, O. (1993). Intraspecific variability of Lacerta (Archaeolacerta) bonnali Lantz, 1927 (Squamata: Sauria: Lacertidae). Herpetozoa, 6: 129-140. Arribas, O. (1994). Una nueva especie de lagartija de los Pirineos Orientales: Lacerta (Archaeolacerta) aurelioi sp. nov. (Reptilia: Lacertidae). Boll. Mus. reg. Sci. nat. Torino, 12 (1): 327-351. Arribas, O. (1997). Morfología, filogenia y biogeografía de las lagartijas de alta montaña de los Pirineos. Tesis doctoral. Universidad Autónoma de Barcelona. 353 pp. Arribas, O. (1998). Osteology of the Pyrenaean Mountain Lizards and comparison with other species of the collective genus Archaeolacerta Mertens, 1921 s.l. from Europe and Asia Minor (Squamata: Lacertidae). Herpetozoa, 11: 47-70. Arribas, O. (1999a). Taxonomic revision of the Iberian “Archaeolacertae” II: Diagnosis, morphology and geographic variation of ‘Lacerta’ aurelioi Arribas, 1994. Herpetozoa, 11: 155-180. Arribas, O. (1999b). Phylogeny and relationships of the mountain lizards of Europe and Near East (Archaeolacerta Mertens, 1921, Sensu Lato) and their relationships among the Eurasian Lacertid Radiation. Russ. J. Herpetol., 6: 1-22. Arribas, O. J. (2007). Istòria Naturau e Evolucion dera Cernalha Aranesa, Iberolacerta <aranica. Conselh Generau d’Aran. 446 pp. Arribas, O. J. (2014). Iberolacerta (Pyrenesaura) aurelioi (Arribas, 1994). Pp. 458-468. En: Salvador, A. (Coordinador). Reptiles, 2ª edición revisada y aumentada. Fauna Ibérica, vol. 10. Ramos, M. A. et al. (Eds.). Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas, Madrid. 1367 pp. Carranza, S., Arnold, E. N., Amat, F. (2004). DNA phylogeny of Lacerta (Iberolacerta) and other lacertine lizards (Reptilia, Lacertidae). Did competition cause long-term mountain restriction?. Systematics and Biodiversity, 2 (1): 57-77. Crochet, P. A., Chaline, O., Surget-Groba, Y., Debain, C., Cheylan, M. (2004). Speciation in mountains: phylogeography and phylogeny of the rock lizards genus Iberolacerta (Reptilia: Lacertidae). Molecular Phylogenetics and Evolution, 30: 860-866. Mayer, W., Arribas, O. (1996). Allozyme differentiation and relationship among the Iberian-Pyrenean Mountain Lizards (Squamata: Sauria: Lacertidae). Herpetozoa, 9 (1/2): 57-61. Mayer, W., Arribas, O. (2003). Phylogenetic relationships of the European lacertid genera Archaeolacerta and Iberolacerta and their relationships to some other 'Archaeolacertae' (sensu lato) from Near East, derived from mitochondrial DNA sequences. J. Zool. Syst. Evol. Research, 41: 157-161. Odierna, G., Aprea, G., Arribas, O., Capriglione, T., Olmo, E. (1996). The karyology of Iberian Rock Lizards. Herpetologica, 52: 542-550.
Oscar Arribas Fecha de publicación: 5-06-2008 Otras contribuciones: 1. Alfredo Salvador. 17-07-2015 Arribas, O. (2015). Lagartija pallaresa – Iberolacerta aurelioi. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Marco, A. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|
|
|
|
|
|