|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: Aran Rock Lizard, identification, description, size, variation.
Sinónimos y combinaciones Lacerta bonnali aranica Arribas, 1993; Lacerta aranica - Arribas, 1997; Archaeolacerta aranica - Arribas, 1997; Iberolacerta aranica - Arribas, 1997.
Origen y evolución La datación y secuencia de diferenciaciones entre las distintas especies de lagartijas pirenaicas, difieren según la aproximación genética empleada. Con un gen de evolución lenta (el gen mitocondrial ARNr 12S), las diferencias son aproximadamente de un 2% entre las tres especies pirenaicas. Otro estudio que tiene en cuenta el gen mitocondrial cyt b (citocromo b) arroja estas diferencias: 6,2% entre I. bonnali e I. aurelioi, 7,4% entre I. aurelioi e I. aranica, y 9,8% entre I. bonnali e I. aranica. Si se toman en conjunto los tres genes (cyt b + 12S ARNr + C-mos; este último, un gen nuclear de evolución muy lenta), las diferencias conservan la misma escala recíproca: 2,5% entre I. bonnali e I. aurelioi, 3,0% entre I. aurelioi e I. aranica, y 3,7% entre I. bonnali e I. aranica. La separación entre las especies pirenaicas estaría entre 3,0 y 5,4 millones de años (3,8-3,9 millones de años de media, según dos estudios diferentes) (Arribas, 1997; Mayer y Arribas, 2003; Carranza et al., 2004; Crochet et al., 2004). El estudio de varios fragmentos concatenados de ADNmt (Arribas, 2007; S. Carranza y O. Arribas, datos inéditos) consistentes en 2.013 bp (cyt b + 12S + 16S), de los que 822 eran variables y 651 informativos de parsimonia, sugiere que aun siendo casi una tricotomía (los procesos separadores debieron actuar simultáneamente en todos los Pirineos), I. aurelioi sería la especie basal, mientras que I. aranica e I. bonnali serían especies hermanas. Esta relación está de acuerdo con la morfología externa (Arribas, 1997, 2000), pero entra en abierto conflicto con los datos, aparentemente más parsimoniosos, de la osteología y cariología (especialmente, el sistema de cromosomas sexuales) (Odierna et al., 1996; Arribas, 1997, 1998, 1999b, 2000), así como la electroforesis de aloenzimas (Mayer y Arribas, 1996), que cuadran más con que I. aranica fuera la especie basal al grupo. Si I. aurelioi fuera la especie más diferenciada, el cromosoma W bibraquial habría aparecido dos veces por una tendencia inherente en todo el grupo a formar cromosomas bibraquiales a partir de monobraquiales por fusiones robertsonianas, pero solo expresada en dos de las especies (I. bonnali e I. aurelioi). Igualmente, la tendencia a perder los procesos del posocular y posfrontal habría estado presente en el grupo y ocurrido de forma independiente dos veces (con la relación expresada, en realidad se trataría de la “recuperación” de esos procesos, primero el del posfrontal en el clado de [I. bonnali + I. aranica], y luego del postocular en el de I. aranica). Todos estos fenómenos repetidos se ajustan poco con el concepto de parsimonia. Otra posibilidad sería que la pérdida de los procesos hubiera ocurrido en algunas de las especies después de la separación de sus táxones hermanos o del linaje principal. Datos preliminares no publicados de Fulgione, Odierna y Arribas (Arribas, 2007), basados en el estudio de cinco microsatélites del ADN de Iberolacerta aranica, I. aurelioi e I. bonnali, muestran ausencia de flujo genético entre ellas y una separación clara de los genotipos comparados. En este estudio inédito de microsatélites, I. bonnali es la especie basal (contrario a los resultados de la genética mitocondrial y la morfología sensu lato). El número medio de alelos resultó más bajo en I. aranica e I. aurelioi que en I. bonnali, lo que está de acuerdo con la menor diversidad genética encontrada en esas especies. Iberolacerta bonnali (de Monte Perdido, Huesca), I. aurelioi (Pica d’Estats, Lérida) e I. aranica (Barrados, Lérida) mostraron valores de heterocigosidad de 0,515, 0,098 y 0,373, respectivamente, significativamente menores que los encontrados en otros lagartos, incluyendo especies insulares, probablemente debido a fuertes cuellos de botella poblacionales y a sus áreas pequeñas y fragmentadas (excepto en I. bonnali). También Bloor (2006) amplificó seis microsatélites y los encontró polimórficos en I. bonnali, cinco en I. aranica y solo tres en I. aurelioi. Las barreras entre estas especies son los grandes valles (baja altura o alternativamente glaciares durante el Pleistoceno) de la Garona (aranica-bonnali), el estrecho margen “en filo de cuchillo” que separa el Mont Vallier del Macizo de Mont-Roig (aranica-aurelioi), y el valle del Noguera Pallaresa (bonnali-aurelioi). En todos los casos está claro que el margen de diferencias entre ellas es muy pequeño, quedaron aisladas “casi a la vez” o sea, en un margen muy estrecho de tiempo, y en general parece que de forma muy ligada al modelado de las cuencas fluviales finiterciarias y a la dinámica de aparición glaciar y sobreexcavación de las altas cuencas del Garona y el Noguera Pallaresa (Arribas, 2014)1.
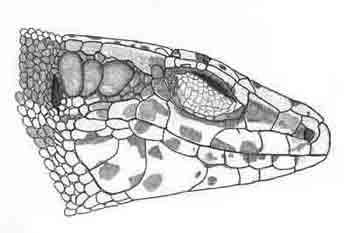
Descripción Folidosis De 0 a 9 (3,3) gránulos supraciliares en el lado derecho y de 0 a 9 (3,54) en el izquierdo; de 18 a 25 (20,98) escamas gulares en los machos y de 18 a 27 (21,53) en las hembras; 7 a 14 (10,69) escamas en el collar; 35 a 48 (39,27) escamas dorsales; 22 a 27 (24,82) escamas ventrales en los machos y 25 a 30 (27,53) en las hembras; 10 a 15 (12,45) poros femorales en el lado derecho y 10 a 16 (12,67) en el izquierdo; 19 a 30 (26,23) laminillas bajo el dedo más largo del pie y de 6 a 11 (7,76) escamas circumanales (Arribas, 2001). Las escamas rostral e internasal están siempre en amplio contacto (100 % de los ejemplares estudiados); la escama postocular y la parietal se encuentran usualmente en contacto (en el 70.6 % de los especímenes estudiados en ambos lados, y en un 9 % sólo en un lado), así como la supranasal y loreal (91.9 % de los especímenes en ambos lados, y en un 6.2 sólo en un sitio). Usualmente sólo existe una escama entre la masetérica y la timpánica (ambas bien visibles), formando las tres un disposición temporal muy característica similar a la que presenta la especie Darevskia mixta del Caucaso (si una disposición temporal similar aparece en algunas I. bonnali, la escama intermedia es usualmente mucho más pequeña que la masetérica y la timpánica) (Figura 1). Alteraciones en las escamas de la cabeza (placas fusionadas, partidas o deformes) son muy frecuentes (Arribas, 2001).
Figura 1. Aspecto lateral de la cabeza de la lagartija aranesa. © O. Arribas.
Coloración Coloración dorsal de tono pardogrisáceo, más o menos claro u oscuro según el color del sustrato que habitan, ocasionalmente con un reflejo olivaceo claro, especialmente en los individuos que han mudado recientemente. Algunos machos presentan el dorso de color verde malaquita (un tono verdeazulado) sobre el grisaceo de fondo (cerca de un 12%). A lo largo del dorso, dos bandas paravertebrales oscuras (80% de los machos y 60% de las hembras) (Figura 2). Pecho y vientre sin pigmento de color, habitualmente en diversos tonos de blanco, ocasionalmente con un reflejo verdoso o azulado que aparece asociado a los tonos malaquita en el dorso y que no es pigmento sino un reflejo (color físico). La región ventral está frecuentemente moteada de negro en los rebordes anteriores de las escamas, especialmente en los machos (90% de los machos y el 30% de las hembras). Pueden existir puntos azules en las escamas ventrales más externas de algunos machos (sólo excepcionalmente y muy pocos en alguna hembra), pero son bastante raros (10%) y parecen estar localizados en zonas muy concretas (Figura 3). Albinismo: Se conocen ejemplares (tres) con zonas despigmentadas en el cuerpo. Usualmente son pequeñas manchas blanco amarillentas, excepto en un ejemplar en el que una banda despigmentada le corria a lo largo de la cola regenerada. Melanismo: Recientemente se han hallado animales muy oscuros (aunque no completamente negros), de un gris pizarra muy oscuro, tanto por el dorso como en el vientre. Ejemplares "concolor": A veces se observan ejemplares anormalmente claros en algún punto concreto. Esos animales presentan el diseño oscuro desaparecido, pero en dos grados, unos completamente desaparecido y otros sólo parcialmente. También se conoce un ejemplar con una fuerte asimetría en el desarrollo de ambos lados del cuerpo, lo que constituye una variación extremadamente rara (Arribas, 2001).
Figura 2. Hembra de lagartija aranesa. © O. Arribas.
Figura 3. Aspecto ventral de la lagartija aranesa. © O. Arribas.
Tamaño Las hembras de I. aranica tienen usualmente mayor talla que los machos (machos: media = 53,84 mm; hembras: media = 56,62 mm). Las diferencias entre ambos sexos son muy significativas. El macho mayor encontrado presentaba una longitud de 61,8 mm desde el extremo del morro hasta la cloaca (ano) y la hembra mayor 66,88 mm. Los miembros anteriores miden de 28.31 a 38.41 % (33.13 %) de la longitud del cuerpo (LCC) en los machos, y de 22.36 % a 38.28 % (30.95 %) en las hembras. El miembro posterior mide desde un 39.32 % a un 53.47 % (45.50 %) de la LCC en los machos, y de un 36.91 % a un 52.41 % (42.31 %) en las hembras. La longitud del píleo (cabeza) representa de 1.82 a 2.83 (1.96) veces su anchura. La placa masetérica mide de un 34.84 a un 57.5 % (46.03 %) de la longitud de la escama parietal, y de un 21.48 % a 55 % (42.1 %) en las hembras. La placa timpánica mide de 0 (no diferenciada) a un 61.53 % (44.96 %) de la longitud de la escama parietal. La placa anal mide de un 40.42 a un 84.71% (60.11 %) en longitud respecto a su anchura en los machos, y de un 45.71 a un 85.86 % (68.98 %) en las hembras. Variación geográfica en la coloración La mayor variación se da en la coloración dorsal, que parece seleccionarse según proporcione mejor o peor camuflaje a los animales en el sustrato rocoso en el que viven. En rocas calcáreas, las lagartijas son frecuentemente muy claras (por ejemplo en la zona de Liat y algunas de la Sierra de Armeros), mientras que sobre pizarras y esquistos son muy oscuras, añadiéndose además el factor del polvillo negro procedente de estas últimas que impregna profundamente (coloración cosmética) a los animales. Sobre filitas (como en Forcall) son también bastante claras. Variaciones de la coloración con la edad Crías recien nacidas (primer año de calendario): dorso gris o pardogrisaceo como en los adultos, usualmente uniforme (ráramente con las dos lineas paravertebrales ligeramente indicadas), y con las dos bandas dorsales bien visibles. Vientre blanco, con un conspicuo moteado negro en el reborde anterior de las escamas ventrales, muy desarrollado en relación con los adultos. Cola usualmente gris, como el dorso (Sª de Armeros, Coll de Barrados) o azulada (Orlà), a veces simplemente blanquecina (diferente del dorso, pero sin desarrollar el pigmento de color). Ejemplares jóvenes y subadultos (hasta su tercer año de calendario incluido) son muy similares al patrón de coloración descrito para las crias recien nacidas, pero sin ningún color vivo presente en la cola, procedan de donde procedan (el color, de estar presente, se pierde durante el segundo año de calendario). El patrón oscuro de las partes ventrales aparece progresivamente menos extendido (cada vez menor respecto a la parte blanca de la escama). Diseño dorsal: las líneas paravertebrales de puntos y las dorsolaterales [=supratemporales] (si están presentes, sólo en la parte más anterior del dorso) aparecen progresivamente en los subadultos (hasta su tercer año de calendario). Adultos (coloración desde el cuarto año de calendario) muy similar al patrón básico de los juveniles, con un diseño ventral oscuro comparativamente menos extendido en cada escama que en éstos (Arribas, 1993, 2001).
Cariotipo Al contrario que las otras dos especies pirenaicas, tanto los machos como las hembras de I. aranica muestran 26 cromosomas: metafases con 10 cromosomas bibraquiales y 16 monobraquiales, similares a los de I. aurelioi en su morfología y meiocitos con 13 pares de homólogos. Contrariamente a las otras dos especies pirenaicas de Iberolacerta (Pyrenesaura), los cromosomas sexuales no son discernibles por técnicas convencionales de tinción. I. aranica debe poseer un sistema ZW de cromosomas sexuales, homomorficos y homocromáticos, ya que se asume que todos los lacértidos presentan heterogametia sexual femenina independientemente de que sea discernible o no citológicamente. El NOR está presente en posición telomérica en el brazo largo del tercer par de cromosomas (tipo-L) como en las otras Iberolacerta (Pyrenesaura). El bandeo C revela intensas bandas centromericas en los cromosomas monobraquiales y débiles bandas centroméricas y pericentroméricas en los bibraquiales. Las bandas centroméricas son completamente digeridas por el encima Alu I, mientras que las pericentroméricas de los tres primeros bibraquiales, la subtelomérica asociada al NOR y la intensa pericentromérica del septimo par (monobraquial) son resistentes a esta tinción. Sólo esta última banda es positiva a la tinción con DAPI. Las dos poblaciones estudiadas (Armeros y Orlà) presentan iguales características en sus cariotipos (Odierna et al., 1996).
Osteología Cráneo con los procesos anterodistal del postfrontal y anteromedial del postorbital presentes y bien desarrollados, lo que la distingue de las dos otras especies pirenaicas. El hueso postorbital es ligeramente más largo o subigual que el postfrontal. La sutura entre el postorbital y el escamoso ocupa cerca de la mitad de la su longitud. Premaxila con siete dientes y un proceso nasal esbelto, de bordes subparalelos (en algún ejemplar aparece algo ensanchado hacia su ápice, que toma un aspecto sublanceolado, incluso con dos o tres prolongaciones en su ápice). De 14 a 17 dientes maxilares (media 15.41) y de 16 a 20 (media 18.58) dientes en el dentario. Dientes bicúspides dominantes sobre los monocúspides. Los machos usualmente tienen 26 vertebras presacrales, y las hembras 27. La tercera vértebra tiene frecuentemente asociada una vértebra osea corta. Fórmula esternal-xifiesternal (3+2). Fontanela esternal oval o redondeada. Claviculas abiertas (marginadas) e interclavicula cruciforme típica (razón entre las ramas anterior y posterior de 0.20 a 0.35, media 0.26). Usualmente seis vertebras dorsales posteriores cortas (un macho joven de Orlà presentaba cinco). La primera vertebra preautotómica caudal con procesos perpendiculares sólo en su parte anterior (tipo-A). (Arribas, 1998).
Referencias Arribas, O. (1993). Intraspecific variability of Lacerta (Archaeolacerta) bonnali Lantz, 1927 (Squamata: Sauria: Lacertidae). Herpetozoa, 6: 129-140. Arribas, O. (1997). Morfología, filogenia y biogeografía de las lagartijas de alta montaña de los Pirineos. Tesis doctoral. Universidad Autónoma de Barcelona.353 pp. Arribas, O. (1998). Osteology of the Pyrenaean Mountain Lizards and comparison with other species of the collective genus Archaeolacerta Mertens, 1921 s.l. from Europe and Asia Minor (Squamata: Lacertidae). Herpetozoa, 11: 47-70. Arribas, O. (1999b). Phylogeny and relationships of the mountain lizards of Europe and Near East (Archaeolacerta Mertens, 1921, Sensu Lato) and their relationships among the Eurasian Lacertid Radiation. Russ. J. Herpetol., 6: 1-22. Arribas, O. (2000). Taxonomic revision of the Iberian “Archaeolacertae” III: Diagnosis, morphology, and geographic variation of Iberolacerta bonnali (Lantz, 1927). Herpetozoa, 13 (3/4): 99-131. Arribas, O. (2001). Taxonomic revision of the Iberian ‘Archaeolacertae’ IV. Diagnosis, morphology and geographic variation of Iberolacerta aranica (Arribas, 1993). Herpetozoa, 14: 31-54. Arribas, O. J. (2007). Istòria Naturau e Evolucion dera Cernalha Aranesa, Iberolacerta aranica. Conselh Generau d’Aran. 446 pp. Arribas, O. J. (2014). Iberolacerta (Pyrenesaura) aranica (Arrbas, 1993). Pp. 444-458. En: Salvador, A. (Coordinador). Reptiles, 2ª edición revisada y aumentada. Fauna Ibérica, vol. 10. Ramos, M. A. et al. (Eds.). Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas, Madrid. 1367 pp. Carranza, S., Arnold, E.N., Amat, F. (2004). DNA phylogeny of Lacerta (Iberolacerta) and other lacertine lizards (Reptilia, Lacertidae). Did competition cause long-term mountain restriction?. Systematics and Biodiversity, 2 (1): 57-77. Crochet, P. A., Chaline, O., Surget-Groba, Y., Debain, C., Cheylan, M. (2004). Speciation in mountains: phylogeography and phylogeny of the rock lizards genus Iberolacerta (Reptilia: Lacertidae). Molecular Phylogenetics and Evolution, 30: 860-866. Mayer, W., Arribas, O. (1996). Allozyme differentiation and relationships among the Iberian-Pyrenean Mountain lizards (Squamata: Sauria: Lacertidae). Herpetozoa, 9 (1/2): 57-61. Mayer, W., Arribas, O. (2003). Phylogenetic relationships of the European lacertid genera Archaeolacerta and Iberolacerta and their relationships to some other 'Archaeolacertae' (sensu lato) from Near East, derived from mitochondrial DNA sequences. J. Zool. Syst. Evol. Research, 41: 157-161. Odierna, G., Aprea, G., Arribas, O., Capriglione, T., Olmo, E. (1996). The karyology of Iberian Rock Lizards. Herpetologica, 52: 542-550.
Oscar Arribas Fecha de publicación: 24-04-2008 Otras contribuciones: 1. Alfredo Salvador 1-09-2015 Arribas, O. (2015). Lagartija aranesa – Iberolacerta aranica. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Marco, A. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|
|
|
|
|
|