|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Northern Raccoon, description, size, variation.
Sistemática La familia Procyonidae (Mammalia: Carnivora) incluía las subfamilias Potosinae (Potos y Bassaricyon; arborícolas y frugívoros) y Procyoninae (Bassariscusy, Nasua y Procyon, más terrestres y omnívoros); una clasificación basada en caracteres morfológicos —dentición, cráneo, esqueleto postcraneal, cola anillada— (Decker y Wozencraft, 1991; Baskin, 2004). Pero la semejanza no se debe a parentesco sino a evolución convergente, por adaptación al hábitat y a la dieta; son grupos ecológicos, no evolutivos (Koepfli et al., 2007; Fulton y Strobeck, 2007). Ahora se reconocen 3 linajes, mediante análisis de caracteres moleculares (Koepfli et al., 2007; Fulton y Strobeck, 2007): los kinkajú (Potos); los olingos y coatíes (Bassaricyon, Nasua); y los gatos de cola anillada y mapaches (Bassariscus, Procyon). El primer prociónido conocido (Pseudobassaris riggsi) se localiza en Europa Occidental y se data en el Oligoceno tardío, aunque la diversificación del grupo ocurre en los bosques tropicales de Centroamérica y el sur de Norteamérica durante el Mioceno (Koepfli et al., 2007; Fulton y Strobeck, 2007). Es probable que el mecanismo de especiación fuera la competencia por recursos, que explicaría la coexistencia de distintos géneros de prociónidos en la misma comunidad (Koepfli et al., 2007). El área de distribución actual de los prociónidos es el continente americano. El panda rojo asiático (Ailurus fulgens), que integra la familia monotípica Ailuridae, antes se definía como otra subfamilia de prociónidos (Fulton y Strobeck, 2007). El género Procyon incluye tres especies: -Mapache común o norteño (Procyon lotor Linnaeus, 1758). En el Centro y Norte de América e introducido en islas caribeñas (las antes descritas como especies endémicas: P. maynardi, Bahamas; P. minor, Guadalupe; y P. gloveralleni, Barbados; Helgen et al., 2008) y del Pacífico (P. insularis, Islas Marías; Helgen y Wilson, 2005), Alaska, Europa y Japón. En expansión a nivel mundial debido a la introducción del mapache por la industria peletera, como especie cinegética y mascota. Especie catalogada como Preocupación menor (Cuarón et al., 2008). -Mapache cangrejero o sureño (Procyon cancrivorus G. [Baron] Cuvier, 1798). En el Centro (en simpatría con el común) y Sur de América, también en Trinidad y Tobago. Especie no amenazada; Preocupación menor (Reid y Helgen, 2008). -Mapache enano o de Cozumel (Procyon pygmaeus Merriam, 1901). Especie endémica de la isla Cozumel (478 km2), en la costa de la península de Yucatán, mar Caribe. De pequeño tamaño (un 20-40% menor que especie continental) y unos 3-4 kg de peso, por adaptación a la escasez de recursos y a la ausencia de depredadores (enanismo insular; McFadden y Meiri, 2013). En peligro crítico, alto riesgo de extinción en estado salvaje (Cuarón et al., 2008).
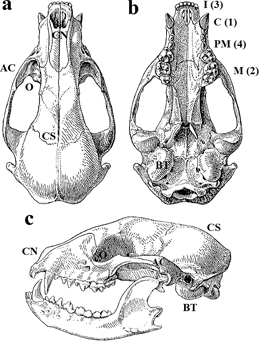
Descripción El mapache es un carnívoro de tamaño mediano. La masa corporal, entre 4 y 12 kg, depende de la latitud (mayor tamaño corporal de los individuos norteños, ajuste a la regla de Bergmann; Meiri et al., 2004), el sexo (los machos son mayores, por selección sexual; Ritke, 1990a) y la estación del año (pérdida de peso en invierno, durante el letargo; Mech et al., 1968). La cabeza es corta y ancha, el cuerpo rechoncho y las patas cortas. Las orejas están separadas y son redondeadas, los ojos son grandes y negros, y el hocico es corto y apuntado. El pelaje es largo y denso, de color gris, negruzco o rojizo, más claro en los flancos y las patas. El albinismo es raro, es un caracter recesivo (Hogan, 1988). Los rasgos distintivos del mapache son la cola anillada, entre 5 y 7 bandas oscuras, y la máscara facial negra a modo de antifaz. El cráneo es compacto y alargado, la región facial es corta y la caja craneal amplia (Figura 1). La cresta sagital, sólo desarrollada en los adultos, no es conspicua. El arco cigomático es robusto. La órbita es grande, la cámara nasal amplia (Figuras 1) y la bulla timpánica redonda y abombada (Stubbe, 1993). La fórmula dentaria del mapache es: 3.1.4.2 / 3.1.4.2, 40 piezas en total. La dentición es primitiva, las cúspides de premolares y molares son redondeadas y aplanadas (bunodonto), propia de un hipocarnívoro, y es frecuente la oligodontia de los primeros premolares (Koepfli et al., 2007).
Figura 1. Vistas dorsal (a), ventral (b) y lateral (c) del cráneo, y oclusal de la serie yugal superior (c). AC: arco cigomático; BT: bulla timpánica; C: canino; CN: cámara nasal; CS: cresta sagital; I: incisivo; M: molar; O: Órbita; PM: premolar. (Modificado de Heptner y Naumov, 1967).
El báculo o hueso peneano es largo y curvo. La función del baculum es incierta; facilitar la penetración, asegurar la fecundación o estimular la ovulación de la hembra (Larivière y Ferguson, 2002). El báculo se usa para determinar la edad de los machos (Sanderson, 1961) o, al menos, distinguir entre jóvenes y adultos por el peso, la longitud y la osificación de la cabeza distal, que en los jóvenes es cartilaginosa (Schwery et al., 2011). La inteligencia del mapache es similar a la de los perros y gatos domésticos y menor que la de los primates, tiene gran capacidad de aprendizaje y los órganos de los sentidos desarrollados; oído y olfato finos, visión nocturna y agudeza del tacto en las manos (Lotze y Anderson, 1979). La palabra mapache deriva de mapachtli, que en lengua náhuatl significa “el que tiene manos”, y el término inglés raccoon del algonquino arahkunem, "el que rasguña con las manos"; en referencia a la habilidad del mapache para manipular objetos. El mapache acostumbra a lavar el alimento antes de comerlo (el epíteto específico lotor significa lavador; también los nombres vernáculos en francés, raton laveur, y alemán, waschbär), aunque es más bien un comportamiento de exploración (Lotze y Anderson, 1979) y una técnica de caza, para localizar y capturar presas bajo el agua (cangrejos y otros invertebrados acuáticos, peces, ranas).
Identificación El mapache puede confundirse con la jineta (Genetta genetta), por la cola anillada, y con el tejón (Meles meles), por la máscara facial y el aspecto general del cuerpo. La cola de la jineta es más larga y tiene entre 8 y 10 anillos, además el pelaje es moteado mientras que el del mapache es uniforme. La cola del tejón es corta, las patas son más robustas y las orejas y los ojos son pequeños. Además, la máscara facial es distinta: la del mapache se extiende alrededor de los ojos y las mejillas, la del tejón la forman dos bandas negras desde el hocico hasta las orejas. Es muy frecuente la confusión entre el mapache y el perro mapache (Nyctereutes procyonoides), un cánido nativo de Asia Oriental y también invasor en Europa, introducido por la industria peletera (Kauhala y Kowalczyk, 2011).
Rastros La observación de mapaches, esquivos y activos de noche, no es frecuente. Los rastros, en cambio, se encuentran sin apenas esfuerzo y la identificación es casi siempre segura (Figura 2). El rastreo es la mejor técnica para detectar al mapache (Salgado, 2012).
Figura 2. Rastros del mapache. a y b. Huellas de la mano y pie izquierdos. c. Pisada de mapache en la orilla del río Jarama. d y e. Letrinas, junto a un tocón y sobre un tronco. f. Restos de cangrejo rojo americano en el mismo lecho del río; el mapache sólo come el abdomen.
Huellas El mapache es un animal plantígrado, apoya la planta del pie para andar (Figura 2). La almohadilla intermedia es semicircular, siempre marca los cinco dedos —largos, finos y separados— y rara vez las uñas —cortas, comprimidas y curvadas–. La huella del pie es mayor cuando imprime el talón. Suele andar al paso, el pie pisa detrás de la mano opuesta.
Excrementos En general, el excremento de mapache es más parecido al de un cánido, grueso y corto, que al de un mustélido, largo y cilíndrico (Figura 2). Cuando come peces y cangrejos el excremento está formado por escamas y espinas y es de color grisáceo o negruzco, o por piezas del caparazón y es rojo intenso. Desprende un olor a pescado o marisco. Es deleznable y blanquea al envejecer. Si se alimenta de frutos e insectos el excremento se compone de las partes duras del fruto y los restos de la cutícula de artrópodos. Los formados por pelo de roedores y conejo pueden confundirse con los de zorro o gato, pero sin el extremo acabado en un penacho de pelo, característico de los cánidos, ni segmentado, propio de los félidos (además, el gato doméstico suele enterrarlo). El mapache usa letrinas, acumula los excrementos sobre troncos caídos y en horquillas de las grandes ramas de los árboles de ribera (Figura 2), incluso en el tejado de casetas y edificios.
Restos de alimento En verano, cuando baja el nivel del agua, acude a la orilla o al propio lecho del río a comer cangrejos. Sólo aprovecha el abdomen, desprecia el cefalotórax (Figura 2). También caza aves acuáticas (pollos y adultos en muda, la “mancada”) y acostumbra a subir la presa a un árbol para devorarla. Sobre las ramas cuelgan los restos, muchas veces el cadáver despechugado. También ataca los nidos de galápago (Álvarez 2008), los excava para comer los huevos. Se reconocen entonces las áreas de desove por los hoyos y las cáscaras de huevo esparcidas.
Guarida Durante el día descansa en cuevas, madrigueras abandonadas o entre marañas de vegetación, e incluso se guarece en casas e infraestructuras rurales y urbanas (Hoffmann y Gottschang, 1977). Las hembras usan los huecos de los grandes árboles como paridera y para la cría de los cachorros (Suzuki, 2003).
Biometría Los machos son mayores que las hembras (Tabla 1). El dimorfismo sexual en tamaño es resultado de la selección sexual, por la competencia entre los machos por el territorio y el acceso a las hembras durante la estación reproductora (Ritke, 1990a). La cola es corta, la mitad de la longitud de cuerpo y cabeza. Las orejas son grandes. En la Tabla 1 se presentan las medidas corporales de machos y hembras de distintas poblaciones de mapache en las áreas de distribución natural e invadida.
Tabla 1. Medidas corporales (media o rango) de individuos adultos de distintas poblaciones de mapache norteamericanas, europeas y japonesas. Masa corporal (kg) y medidas morfométricas; mm): LT: longitud total, LCU: longitud del cuerpo, y LCO: longitud de la cola. *Las medidas corporales, de mapaches capturados, fueron tomadas en centros de recuperación de fauna salvaje (CRAS Madrid, CRFS Guadalajara, CAAE Baleares) y los datos cedidos por las Consejerías de Medio Ambiente y Ordenación del Territorio de la Comunidad de Madrid, de Agricultura de Castilla-La Mancha y de Agricultura, Medio Ambiente y Territorio de las Islas Baleares.
Voz Los mapaches emiten un sonido largo y trémulo, un quejumbroso whoo-oo-oo-oo, parecido al ulular de los búhos (Murie y Elbroch, 2005). El repertorio vocal del mapache es rico; llamadas de socorro a miembros del grupo, voces de amenaza para repeler a otros animales y de aproximación para establecer contacto con otros individuos (Sieber, 1984).
Variación geográfica En el área de distribución natural, los mapaches norteños son más grandes que los del sur (Meiri et al., 2004; Tabla 1); un patrón geográfico también longitudinal, la talla varía de noroeste a sureste de Norteamérica (Ritke y Kennedy, 1988). Las medidas de cuerpo y cráneo de los mapaches establecidos en Europa corresponden a la especie de Norteamérica (Tabla 1). En el área de introducción principal, en Hesse (Alemania), es frecuente la ausencia de los primeros premolares (Lutz, 1995); por efecto fundador; la población deriva de unos pocos individuos (Frantz et al., 2013). En general, la baja diversidad génica y los efectos de cuello de botella es resultado del proceso de fundación de las poblaciones de mapache europeas, casi siempre por unos pocos individuos liberados o escapados (Biedrzycka et al., 2013). Nuevas introducciones o la conexión entre poblaciones en expansión incrementaría la variabilidad genética por flujo génico y, por tanto, la viabilidad de la especie en el ecosistema receptor. En el interior de la península ibérica, entre 2 y 4 individuos fundan las subpoblaciones de los ríos Jarama y Henares, que están ahora en contacto (Alda et al. 2012).
Muda El largo, suave y denso pelaje de invierno es protector, impermeable y aislante térmico. En primavera lo muda y el pelo es entonces más corto y fino (Goldman, 1950). Debido a la densidad y la textura del pelaje invernal, la piel de mapache es apreciada en peletería para la confección de prendas de abrigo.
Referencias Alda, F., Ruiz-López, M. J., García, F. J., Gompper, M. E., Eggert, L. S., García, J. T. (2013). Genetic evidence for multiple introduction events of raccoons (Procyon lotor) in Spain. Biological Invasions, 15(3): 687-698. Álvarez, A. (2008). Predación de nidos de galápago leproso (Mauremys leprosa) por mapaches. Quercus, 269: 49. Banfield, A. W. F. (1974). Mammals of Canada. University of Toronto Press. Toronto, Ontario, Canadá. 438 pp. Bartoszewicz, M., Okarma, H., Zalewski, A., Szczesna, J. (2008). Ecology of the raccoon from western Poland. Annales Zoologici Fennici, 45(4): 291-298. Baskin, J. A. (2004). Bassariscus and Probassariscus (Mammalia, Carnivora, Procyonidae) from the early Barstovian (Middle Miocene). Journal of Vertebrate Paleontology, 24(3), 709-720. Biedrzycka, A., Zalewski, A., Bartoszewicz, M., Okarma, H., Jędrzejewska, E. (2014). The genetic structure of raccoon introduced in Central Europe reflects multiple invasion pathways. Biological Invasions, 16(8): 1611-1625. Cuarón, A. D., de Grammont, P. C., Vázquez-Domínguez, E., Valenzuela-Galván, D., García-Vasco, D., Reid, F., Helgen, K. (2008a). Procyon pygmaeus. En: UICN 2009. UICN Lista Roja de Especies Amenazadas. Cuarón, A. D., Reid, F., Helgen, K. (2008b). Procyon lotor. En: UICN 2014. UICN Lista Roja de Especies Amenazadas. Decker, D. M., Wozencraft, W. C. (1991). Phylogenetic analysis of recent procyonid genera. Journal of Mammalogy, 72(1), 42-55. Frantz, A. C., Heddergott, M., Lang, J., Schulze, C., Ansorge, H., Runge, M. (2013). Limited mitochondrial DNA diversity is indicative of a small number of founders of the German raccoon (Procyon lotor) population. European Journal of Wildlife Research, 59(5): 665-674. Fulton, T. L., Strobeck, C. (2007). Novel phylogeny of the raccoon family (Procyonidae: Carnivora) based on nuclear and mitochondrial DNA evidence. Molecular Phylogenetics and Evolution, 43(3): 1171-1177. Goldman, E. A. (1950). Raccoons of North and Middle America. North American Fauna, 60: 1-153. U.S. Fish and Wildlife Service. Washington, D.C. Hall, E. R. y Kelson, K. R. (1959). The mammals of North America. Ronald Press Company. New York, EE.UU. 1083 pp. Helgen, K. M., Maldonado, J. E., Wilson, D. E., Buckner, S. D. (2008). Molecular Confirmation of the Origin and Invasive Status of West Indian Raccoons. Journal of Mammalogy, 89(2): 282-291. Helgen, K. M., Wilson, D. E. (2005). A systematic and zoogeographic overview of the raccoons of Mexico and Central America. En: Sanchez-Cordero V., Medellin R. A. (eds.). Contribuciones Mastozoologicas: en Homenaje a Bernardo Villa. Pp. 221-236. Instituto de Biología e Instituto de Ecología, UNAM, México. Heptner, V. G., Naumov, N. P., Yurgesson, P. B., Sludsky, A. A., Chirkova, A. F., Bannikov, A. G. (1967). Mammals of the USSR. volumen II, parte 1b. Moscú. Hoffmann, C. O., Gottschang, J. L. (1977). Numbers, distribution and movements of a raccoon population in a suburban residential community. Journal of Mammalogy, 58(4): 623-636. Hogan, A. (1988). Two independent loci for albinism in raccoons Procyon lotor. Journal of Heredity, 79(5): 387-389.
Kauhala, K., Kowalczyk, R. (2011). Invasion of the raccoon dog Nyctereutes procyonoides in Europe: History of colonization, features behind its success, and threats to native fauna —a review. Current Zoology, 57(5): 584-598. Koepfli, K.-P., Gompper, M. E., Eizirik, E., Ho, C.-C., Linden, L., Maldonado, J. E., Wayne., R. K. (2007). Phylogeny of the Procyonidae (Mammalia: Carnivora): molecules, morphology and the Great American Interchange. Molecular Phylogenetics and Evolution, 43(3): 1076-1095. Larivière, S., Ferguson, S. H. (2002). On the evolution of the mammalian baculum: vaginal friction, prolonged intromission or induced ovulation? Mammal Review, 32(4): 283-294. Lotze, J. H., Anderson, S. (1979). Procyon lotor. Mammalian Species, 119: 1-8. Lutz, W. (1995). Occurrence and morphometrics of the racoon Procyon lotor L. in Germany. Annales Zoologici Fennici, 32(1): 15-20. McFadden, K. W., Meiri, S. (2013). Dwarfism in insular carnivores: a case study of the pygmy raccoon. Journal of Zoology, 289(3): 213-221. Mech, L. D., Barnes, D. M., Tester, J. R. (1968). Seasonal weight changes, mortality, and population structure of raccoons m Minnesota, Journal of Mammalogy, 49(1): 63-73. Meiri, S., Dayan, T., Simberloff, D. (2004). Carnivores, biases and Bergmann’s rule. Biological Journal of the Linnean Society, 81(4): 579-588. Murie, O. J. (1974). Animals Track. Peterson Field Guild Series (2ª edición). Houghton Mifflin, Boston. 375 pp. Peterson, R. L. (1966). The mammals of Eastern Canada. Oxford University Press. Toronto, Ontario, Canadá. 465 pp. Reid, F., Helgen, K. (2008). Procyon cancrivorus. En: IUCN 2013. UICN Lista Roja de Especies Amenazadas. Ritke, M. E. (1990a). Sexual dimorphism in the raccoon Procyon lotor: morphological evidence for intrasexual selection. American Midland Naturalist, 124(2): 342-351. Ritke, M. E., Kennedy, M. L. (1988). Intraspecific morphological variation in the raccoon (Procyon lotor) and its relationship to selected environmental variables. Southwestern Naturalist, 33(3): 295-314. Salgado, I. (2012). Guía de identificación de los rastros del mapache común (Procyon lotor): aplicación de las técnicas de rastreo a la detección de especies exóticas invasoras. (2012) En: GEIB Grupo Especialista en Invasiones Biológicas (ed.). EEI 2012 Notas Científicas. Pp. 213-215. 4º Congreso Nacional sobre Especies Exóticas Invasoras "EEI 2012". GEIB, Serie Técnica Nº 5. León, 218 pp. Sanderson, G. C. (1961). Techniques for determining age of raccoons. Illinois Natural History Survey, Biological Notes, 45: 1-16. Schwery, O., Köhnemann, B. A., Michler, F.-U., Brinkmann, W. (2011). Morphometrical characterization of a raccoon (Procyon lotor L.) population from Müritz National Park (Germany) by means of the Os baculum. Beiträge zur Jagd- und Wildforschung, 36: 605-617. Sieber, O. J. (1984). Vocal communication in raccoons (Procyon lotor). Behaviour, 90(1-3): 80-113. Stubbe, M. (1993). Procyon lotor (Linné, 1758) – Waschbär. Pp. 331-364. En: Stubbe, M., Krapp, F. (Eds.). Band 5: Raubsäuger – Carnivora (Fissipedia). Teil I: Canidae, Ursidae, Procyonidae, Mustelidae 1. En: Niethammer, J., Krapp, F. (Eds.). Handbuch der Säugetiere Europas. Aula-Verlag, Wiesbaden. Suzuki, T., Toshiki, A., Maekawa, K. (2003). Spacing pattern of introduced female raccoons (Procyon lotor) in Hokkaido, Japan. Mammal Study, 28(2): 121-128.
Iván Salgado García Fecha de publicación: 27-02-2015 Salgado, I. (2015). Mapache – Procyon lotor. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Barja, I. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||