|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Schreiber's Long-fingered Bat, description, size, variation.
Sistemática Hay dos clados dentro del género Miniopterus, uno africano, que incluye la especie europea, y otro que contiene las especies de Asia y Australia (Miller-Butterworth et al., 2005). Considerada durante mucho tiempo como una especie cosmopolita, M. schreibersii representa un complejo de especies (Appleton et al., 2004). Actualmente se restringe M. schreibersii a las poblaciones de Europa, norte de África y regiones litorales de Turquía. El taxón pallidus, considerado hasta ahora como una subespecie de M. schreibersii, se ha propuesto separarlo como especie diferente (Furman et al., 2010c).
Descripción Murciélago de tamaño medio, con un hocico muy corto, achatado y con la frente alta y protuberante. Las orejas son cortas y triangulares, con un trago corto, redondeado y proyectado hacia delante. Las alas son estrechas y largas, la segunda falange del tercer dedo es tres veces más larga que la primera y normalmente se encuentra plegada dentro del ala cuando el animal reposa. El plagiopatagio se inserta a la altura del tobillo y el uropatagio carece de lóbulo postcalcáneo. Pelaje corto y muy denso, de color grisáceo en la parte dorsal y más blanquecino en la parte ventral (Cabrera, 1914; Boye, 2004). Fórmula dentaria: 2.1.2.3/3.1.3.3 (Cabrera, 1914). Dotación cromosómica (2n) = 46 (Pérez-Suárez et al., 1991).
Biometría y masa corporal Esta especie tiene unas alas muy largas (longitud tercer dedo; D3) y estrechas (longitud quinto dedo; D5). Las medidas biométricas de los individuos adultos capturados en la Región de Murcia (Tabla 1) (Lisón et al., 2010, 2011, Lisón 2012) revelan que los individuos del sureste son ligeramente más pequeños que los del norte de España y además se observa la existencia de dimorfismo sexual inverso en las medidas alares y que las hembras son ligeramente más grandes que los machos (Lisón, 2012).
Tabla 1. Medidas biométricas (media ± SD) y su rango para la población de M. schreibersii en la Región de Murcia. Abreviaturas: FA+ = longitud del antebrazo incluyendo la muñeca; D5 = longitud del quinto dedo excluyendo la muñeca; D3 = longitud del tercer dedo excluyendo la muñeca. (F. Lisón, datos no publicados).
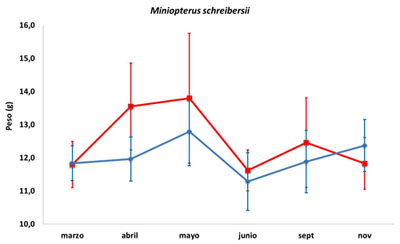
Hay estacionalidad en la masa corporal (Serra-Cobo et al., 2000), tanto para los machos como para las hembras (Figura 1; F. Lisón, datos no publicados). Los machos suelen adquirir una mayor masa corporal a finales de la primavera, perdiendo masa corporal durante los meses más secos. Posteriormente van recuperándose durante el otoño. En las hembras las variaciones son más acusadas, tienen una elevada masa corporal en primavera (especialmente por encontrarse preñadas) y al igual que los machos, pierden masa corporal durante los meses secos del verano, recuperándola durante el otoño (Figura 1).
Figura 1. Variación anual de la masa corporal (media ± SD) en ambos sexos de Miniopterus schreibersii para la Región de Murcia (F. Lisón, datos no publicados). La línea azul indica la masa corporal de los machos y la línea roja la masa corporal de las hembras.
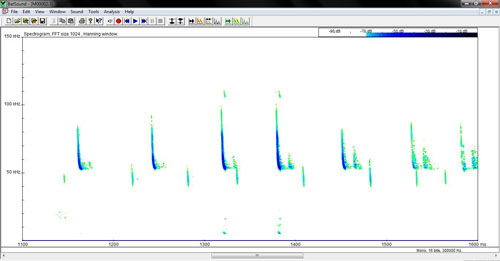
Ultrasonidos Emite ultrasonidos tipo FM-QCF (Frecuencia modulada-cuasi frecuencia constante; Dietz et al. 2009), con una frecuencia de máxima energía alrededor de 54-56 kHz (ver Tabla 2 y Figura 2). Estas llamadas pueden confundirse con las que emite Pipistrellus pygmaeus y deben realizarse análisis estadísticos para su correcta identificación. Los pulsos de P. pygmaeus suelen ser irregulares.
Tabla 2. Valores de los parámetros de las llamadas de ecolocación de Miniopterus schreibersii en diferentes áreas de Europa. Se indica la media ± SD. Los valores para las frecuencias se dan en kilohertzios (kHz) y los valores de tiempo se dan en milisegundos (ms). Acrónimos: Fi = Frecuencia inicial; Ff = Frecuencia final; FMaxE = Frecuencia de máxima energía; D = Duración; IPI = Intervalo entre pulsos.
Figura 2. Espectrograma de los pulsos de ecolocación de Miniopterus schreibersii (F. Lisón, datos no publicados).
Variación geográfica Muestra escasa diferenciación genética entre los poblaciones orientales y occidentales, sugiriéndose que podría haber recolonizado Europa a partir de un refugio glacial (Furman et al., 2010a). No se han observado diferencias genéticas entre las poblaciones de las islas Baleares y de Cataluña (Amengual et al., 2007). Las poblaciones de Pirineos tiene coloración más clara y parda que las de Austria (Bauer, 1956). La diversidad de haplotipos es la mitad en el norte de Portugal en comparación con el centro y el sur (Ramos Pereira et al., 2009).
Referencias Amengual, B., López-Roig, M., Mas, O., González, J., Serra-Cobo, J. (2007). Analisi d' ADN mitocondrial de cinc especies de quiropters de les Illes Balears. Bolleti de la Societat d'Historia Natural de les Balears, 50: 269-277. Appleton, B. R., McKenzie, J. A., Christidis, L. (2004). Molecular systematics and biogeography of the bent-wing bat complex Miniopterus schreibersii (Kuhl, 1817) (Chiroptera: Vespertilionidae). Molecular Phylogenetics and Evolution, 31 (2): 431-439. Bauer, K. (1956). Zur Kentnis der Fledermausfauna Spaniens. Bonner Zoologische Beitrage, 7: 296-319. Boye, P. (2004). Miniopterus schreibersii Natterer in Kuhl, 1819 – Longflügelfledermaus. Pp. 1093-1122. En: Krapp, F. (Ed.). Band 4: Fledertiere. Teil II: Chiroptera II Verpertilionidae 2, Molossidae, Nycteridae. En: Niethammer, F., Krapp, F. (Eds.). Handbuch der Säugetiere Europas. Aula Verlag, Wiebelsheim. Cabrera, A. (1914). Fauna Ibérica. Mamíferos. Museo Nacional de Ciencias Naturales, Madrid. Furman, A, Ŏztunç, T., Postawa, T., Çoraman, E. (2010a). Shallow genetic differentiation in Miniopterus schreibersii (Chiroptera: Vespertilionidae) indicates a relatively recent re-colonization of Europe from a single glacial refugium. Acta Chiropterologica, 12(1): 51-59. Furman, A., Postawa, T., Ŏztunç, T., Çoraman, E. (2010c). Cryptic diversity of the bent-wing bat, Miniopterus schreibersii (Chiroptera: Vespertilionidae), in Asia Minor. BMC Evolutionary Biology, 10: 121. Lisón, F. (2012). Datos biométricos de cinco especies de murciélagos (Mammalia: Chiroptera) de la Región de Murcia (SE España). Anales de Biología, 34: 37-42. Lisón, F., Aledo, E. Calvo, J. F. (2011). Los murciélagos (Mammalia: Chiroptera) de la Región de Murcia (SE España): distribución y estado de conservación. Anales de Biología, 33: 79-92. Lisón, F., Yelo, N. D., Haz, Á., Calvo, J. F. (2010). Contribución al conocimiento de la fauna quiropterológica de la Región de Murcia. Galemys, 12(1): 3-20. Miller-Butterworth, C. M., Eick, G., Jacobs, D. S., Schoeman, M. C., Harley, E. H. (2005). Genetic and phenotypic differences between South African long-fingered bats, with a global miniopterine phylogeny. Journal of Mammalogy, 86 (6): 1121-1135. Obrist, M. K., Boesch, R., Flückiger, P. F. (2004). Variability in echolocation call design of 26 Swiss bat species: consequences, limits and options for automated field identification with a synergetic pattern recognition approach. Mammalia, 68 (4): 307-322. Papadatou, E., Butlin, R. K., Altringham, J. D. (2008). Identification of bat species in Greece from their echolocation calls. Acta Chiropterologica, 10: 127-134. Pérez-Suárez, G., Reina, J. M., Navlet, J., De Paz, O., López-Caballero, E. J. (1991). Citogenética de quirópteros en la península ibérica. Pp. 305-330. En: Benzal, J., De Paz, O. (Eds.). Los murciélagos de España y Portugal. Colección técnica. Ministerio de Agricultura, Pesca y Alimentación, Instituto Nacional para la Conservación de la Naturaleza, Madrid. Ramos Pereira, M. J., Salgueiro, P., Rodrigues, L., Coelho, M. M., Palmeirim, J. M. (2009). Population structure of a cave-dwelling bat, Miniopterus schreibersii: does it reflect history and social organization? Journal of Heredity, 100 (5): 533-544. Russo, D., Jones, G. (2002). Identification of twenty-two bat species (Mammalia: Chiroptera) from Italy by analysis of time-expanded recordings of echolocation calls. Journal of Zoology, 258: 91-103. Serra-Cobo, J., López-Roig, M., Marques-Bonet, T., Martínez-Rica, J. P. (2000). Body condition changes of Miniopterus schreibersii in autumn and winter. Revue d'Ecologie la Terre et la Vie, 55 (4): 351-360.
Fulgencio Lisón Fecha de publicación: 22-05-2012 Revisiones: 19-09-2014 Lisón, F. (2017). Murciélago de cueva – Miniopterus schreibersi. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Barja, I.(Eds). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||