|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: European Free-tailed Bat, activity, movements, behaviour.
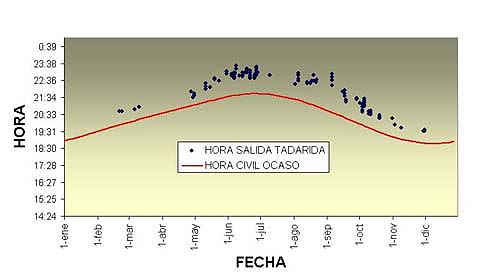
Actividad En Valladolid los rabudos salen más tarde que Pipistrellus pipistrellus y Eptesicus serotinus (especies con las que comparten refugio). Los machos, en época de celo, salen y entran continuamente durante la noche para defender el territorio. Normalmente todos los ejemplares de la colonia regresan antes de las primeras luces del alba (A. Balmori, obs. pers.). A partir de octubre y durante todo el invierno las salidas se adelantan respecto al ocaso, pudiendo observarse algunos ejemplares volando con la luz crepuscular en esta época (Figura 1). Por el contrario Arlettaz (1990) observa que en Los Alpes suizos las salidas más tardías se producen más frecuentemente en enero y febrero. En Portugal , en otoño, la mayoría de los ejemplares deja el refugio al mismo tiempo pero el momento de regreso es muy variable (Marques et al., 2004). ). La disponibilidad de insectos (especialmente de polillas, su principal alimento) muestra un pico después del crepúsculo, pero se mantiene bastante alta durante la noche, haciendo innecesario salir a comer temprano (Marques et al., 2004). En el sur de la Península los patrones de actividad de vuelo varían a lo largo del año. Entre los meses de noviembre a marzo entre un 80-90% de la actividad se restringe a las 3-5 primeras horas de la noche. Durante el resto del año siguen un patrón bimodal con máximos tras el ocaso y antes del orto con actividad variable entre ambos (Ibáñez y Pérez-Jordá, 2004). En Portugal en otoño la mayoría de los murciélagos mostraron un patrón de actividad unimodal, con el máximo aproximadamente tres horas después de la puesta del sol. La salida se produce una hora después de la puesta del sol, con un solo turno de vuelo muy largo, sin descanso, de 6 h. 39 minutos de media. Entre las 20 y las 24 horas casi todos los murciélagos están fuera del refugio, después la proporción de individuos con actividad alimenticia se va reduciendo a lo largo de la noche. La duración no está relacionada con las condiciones meteorológicas sino más bien con el éxito de caza (Marques et al., 2004).
Figura 1. Horario de salida de los refugios de Tadarida teniotis en Valladolid. (Para la representación se ha seguido el horario de verano, dos horas adelantado respecto al solar). (A. Balmori, datos propios).
Es capaz de estar activo con factores meteorológicos adversos como frío, lluvia y viento (Ibáñez y Pérez-Jordá, 2004; A. Balmori, datos propios). En Suiza, por debajo de los 0º no se ha encontrado actividad, entre 0 y 5 º existe actividad de vuelo y a partir de los 5º actividad de caza (Arlettaz, 1990). La actividad intermitente de caza en invierno coincide aparentemente con periodos de dulcificación de la temperatura (Arlettaz, 1990; A. Balmori, obs. pers.). En Portugal durante las noches más frías permanecen en el refugio sin salir. Ni la temperatura ni el viento parecen tener influencia en la duración de los periodos de caza. En noches algo frías e incluso con fuertes vientos salen a comer, pero en tiempo tormentoso retornan al refugio (Marques et al., 2004). El rápido retorno a los refugios con los chubascos tormentosos también se ha observado en Valladolid (Balmori, obs. pers.). Su regulación térmica es deficiente (Arlettaz, 1990; Arlettaz et al., 2000). En los Alpes hiberna en periodos que pueden superar los 8 días. La temperatura corporal media (10-13º C) y la media de frecuencia de actividad invernal (Cada 3,4 días en enero) son marcadamente más altos que en otras especies de zonas templadas (Arlettaz et al., 2000). Es la primera especie de molósido en la que se demuestra una cierta capacidad de hibernación (Arlettaz et al., 2000). Las frecuentes fases de vigilia conducen a un rápido gasto de reservas. Precisamente a la necesidad de reponerlas puede ser achacada la caza en invierno (Arlettaz et al, 2000).
Dominio vital El área de campeo es muy grande, desplazándose a varios kilómetros para alimentarse (Arlettaz, 1990, Fernández, 2002; A. Balmori, obs. pers.). En Suiza se ha estimado entre 6 y 10-15 Km. la distancia entre los cazaderos y los refugios (Arlettaz, 1990). En Valladolid se han detectado ejemplares cazando a unos 14 Km del refugio más próximo (A. Balmori, datos propios). Al amanecer y anochecer son capaces de volar largos tramos sin utilizar ecolocación (Arlettaz, 1990).
Movimientos Existe cierta discrepancia sobre sus movimientos estacionales. Algunos autores han propuesto que se trata de una especie migratoria (Saint Girons, 1973) mientras otros la consideran sedentaria (Arlettaz, 1990). Los refugios estudiados en España (Huelva y Valladolid) muestran una caída del número de ejemplares en invierno (Balmori, 2003a; Ibáñez y Pérez-Jordá, 2004), por lo que podría tratarse de una especie parcialmente migradora. En una colonia que utiliza las juntas de dilatación de un puente en Villarrasa (Huelva), los murciélagos ocupan el refugio la mayor parte del año aunque el mayor número de individuos se observa durante la primavera (50-60) y el otoño (80) y la mínima en verano (10) e invierno (0-20) (Ibáñez y Pérez-Jordá, 1998). En Valladolid desaparece la mayoría de la colonia en invierno quedando solamente unos pocos machos (Balmori, 2003a). Lo mismo ocurre en una colonia estudiada en Francia que se ocupa entre marzo y abril y se abandona en noviembre, aunque algunos ejemplares están presentes antes de Navidad. La colonia estudiada en Portugal permanece en invierno, pero con menor número de individuos. A finales de primavera y en verano la colonia también es menor (Marques, com. pers.). Estos datos hacen suponer ciertos movimientos (Mille, 1988). En el Alto Aragón apenas existen citas del murciélago rabudo en invierno. Las observaciones son casi todas entre abril y octubre (Woutersen y Bafalui Zoriguel, 2001).
Patrón social y comportamiento Se han obtenido evidencias de una organización social compleja. El seguimiento de una colonia de Valladolid (Castilla y León, España) mediante observación nocturna y anillamiento ha demostrado la existencia de harenes, constituidos por un macho y un pequeño grupo de hembras (entre 1 y 9 con 2,5 hembras de media por cada macho). Los machos reaccionan agresivamente a la presencia de competidores en su territorio, realizando veloces persecuciones aéreas (acrobáticas en algunos casos) en la proximidad de los refugios, que suelen ser por parejas, pero en las que pueden intervenir hasta 4 o 5 machos (Balmori, 2003a). Estas persecuciones se producen generalmente a una altura de 15 a 30 m sobre el suelo. Son vuelos acrobáticos que realizan a gran velocidad describiendo círculos y cambiando rápidamente de dirección, ciñéndose a los edificios circundantes. Suelen ir acompañados de fuertes emisiones vocales perfectamente audibles (A. Balmori, obs. pers). A veces también planean brevemente con las alas extendidas (Arlettaz, 1990; A. Balmori, obs. pers.). En el estudio realizado en Portugal existen importantes picos de actividad alrededor de los refugios especialmente a la 1 de la madrugada y justo antes de la salida del sol. (Marques et al., 2004), que se corresponderían con los vuelos territoriales descritos. Los machos desprenden un fuerte olor que recuerda al apio (Apium graveolens), utilizado posiblemente para marcar el territorio. En primavera y otoño los machos dominantes emiten llamadas sociales desde los refugios para atraer a las hembras. El comportamiento reproductor se ajusta al modelo "poliginia de defensa de recurso" (Balmori, 2003a). Las conspicuas manchas de orina a la entrada de los refugios revela su exacta localización (Arlettaz, 1993). Es una especie que no requiere termorregulación social por lo que evita el contacto intraespecífico, y no se asocia en grupos, que solo se han observado en invierno (Arlettaz, 2000). Cuando un congénere se aproxima a otro, ambos comienzan a emitir fuertes vocalizaciones (Arlettaz, 1993). Esas interacciones pueden escucharse perfectamente desde el exterior (A. Balmori, obs. pers.)
Referencias Arlettaz, R. (1990). Contribution à l´éco-éthologie du molosse de cestoni, Tadarida teniotis (Chiroptera), dans les Alpes Valaisannes (sud-ouest de la Suisse). Z. Säugetierkunde, 55: 28-42. Arlettaz, R. (1993). Tadarida teniotis´tail. Myotis, 31: 155-162. Arlettaz, R., Ruchet, C., Aeschimann, J., Brun, E., Genoud, M., Vogel, P. (2000). Physiological traits affecting the distribution and wintering strategy of the bat Tadarida teniotis.Ecology, 81: 1004-1014. Balmori, A. (2003a). Avances en el conocimiento de la biología y organización social del murciélago rabudo (Tadarida teniotis). Galemys, 15 (nº especial): 37-53. Fernández, J. (2002). Los murciélagos en Castilla y León. Atlas de distribución y tamaño de las poblaciones. Junta de Castilla y León, Valladolid. Ibáñez, C., Pérez-Jordá, J. L. (1998). Longevity in the European free-tailed bat (Tadarida teniotis). Journal of Zoology, London, 245: 213-214. Ibáñez, C., Pérez-Jordá, J.L. (2004). Tadarida teniotis (Rafinesque, 1814)- Europäische Bulldoggfledermaus. Pp. 1125-1143. En: Krapp, F. (Ed.). Handbuch der Säugetiere Europas.Vol. 4/2. Aula Verlag, Wiesbaden. Marques, J. T., Rainho, A., Carapuco, M., Oliveira, P., Palmeirim, J. M.. (2004). Foraging behaviour and habitat use by the European free-tailed bat Tadarida teniotis. Acta Chiropterologica, 6: 99-110. Mille, J. L. (1988). Une Colonie de Molosses de Cestoni Tadarida teniotis a Sisteron. Faune de Provence, 9: 102-103. Mitchell-Jones, A. J., Amori, G., Bogdanowicz, W., Krystufek, B., Reijnders, P. J. H., Spitzenberger, F., Strubbe, M., Thissen, J. B. M., Vohralík, V., Zima, J. (Eds.) (1999). Atlas of European Mammals. Societas Europaea Mammalogica. T & AD Poyser, London. Saint Girons, M. C. (1973).Les mammifères de France et du Benelux (faune marine exceptée). Doin, Paris. 481 pp. Woutersen, K., Bafalui Zoriguel, J. J. (2001). Murciélagos del Alto Aragón. K. Woutersen Publ., Huesca. 143 pp.
Alfonso Balmori Martínez Fecha de publicación: 26-10-2004 Revisiones: 6-04-2005 Balmori, A. (2017). Murciélago rabudo – Tadarida teniotis. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Barja, I. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|
|
|
|
|
|