|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: Pyrenean Chamois, Cantabrian Chamois, activity, home range, movements, behaviour.
Actividad Ver detalles en Ecología trófica.
Dominio vital Pocos estudios se han realizado hasta el momento destinados a establecer el área de campeo del rebeco pirenaico. Crampe et al. (2004) establecen el área de campeo de las hembras en 20,4 ha en verano y 11,2 ha en invierno en una población no cazada y estabilizada (crecimiento poblacional del 2%) del sector de Cauterets en el P. N. des Pyrénées. Mientras que Levet et al. (1995) estiman un área de campeo para las hembras entre 100 y 200 ha en una población reintroducida a baja altitud. Loison et al. (1999) calculan áreas de campeo de 150 y 300 ha, para machos y hembras respectivamente, en la Reserva d’Orlu, una población de alta densidad (34 indiv/km2) y crecimiento poblacional del 13%. Seguimientos realizados recientemente en Andorra proporcionan cifras de 211 y 190 ha para machos y hembras respectivamente (Solá, 2004).
Movimientos Ver detalles en Ecología trófica. Se ha observado en el Parque Nacional de los Pirineos que todas las hembras de una población se concentran en una zona de refugio. En verano, una parte de las hembras son sedentarias (52,2%) y permanecen en la misma zona de refugio invernal, mientras que otras (46,8%) se desplazan a una distancia horizontal de 1,4-9,2 km y una elevación de 300-800 m superior (Crampe et al., 2007).
Patrón social y comportamiento Cordillera Cantábrica En el rebeco cantábrico se han detectado 4 tipos de grupos sociales, hembras con crías (mediana = 2 – 33, media de medianas = 4), hembras sin crías (mediana = 1 – 5, media de medianas = 1), grupos mixtos (mediana = 2 – 55, media de medianas = 6,5), y machos (mediana = 1 – 16, media de medianas = 8) (Pérez-Barbería y Nores, 1994). Los tamaños de los grupos que no contienen crías son más estables en el tiempo. Los grupos de hembras sin crías nunca excedían de 5 individuos, mientras que los grupos de machos suelen formar agregaciones de hasta 16 individuos, aunque los individuos solitarios también son frecuentes. Los grupos de hembras con crías muestran gran variación en el tamaño de grupo durante la mayor parte del año, la mayor estabilidad tiene lugar durante otoño y primavera. Los grupos de hembras con crías son de mayor tamaño en las áreas desprovistas de terreno de escape. En las zonas con terrenos de escape (riscos, quebradas, grandes pendientes y proximidad al bosque) los rebecos se desplazan con agilidad, detectan la proximidad de los depredadores y rara vez son molestados por excursionistas o ganaderos. Algunos autores han sugerido que la agregación en áreas abiertas desprovistas de terreno de escape está favorecida por el incremento del contacto visual entre miembros del grupo (Hirth, 1977; Jordá, 1981; Elsner-Schack, 1985; Lagory, 1986; Lovari y Consentino, 1986; Richard-Hansen et al., 1992). El incremento de tamaño de grupo de las agregaciones de hembras con crías en terrenos de escape no es una buena estrategia para evitar la depredación. En estas zonas un incremento en el tamaño de grupo incrementaría la localización visual por los depredadores e impediría una huida eficiente. Al comienzo de los partos el tamaño de grupo de las hembras con crías tenía una mediana de 3 a últimos de abril, y de 5,5 en junio. En verano en las zonas desprovistas de terrenos de escape el tamaño de grupo de las hembras con crías incrementaba (mediana = 12), por estas fechas las crías ya se desenvuelven con agilidad y la probabilidad de escapar de un ataque de los depredadores incrementa. En general las zonas desprovistas de terrenos de escape tienen pasto más abundante y de mejor calidad que las zonas de escape (Pérez-Barbería y Nores, 1994). Otra variable que debe ser considerada a la hora de analizar el tamaño de grupo es la densidad poblacional. Utilizando censos de 24 áreas de la montaña cantábrica con presencia de rebeco se obtuvo una correlación positiva entre el tamaño de grupo y la densidad poblacional (mediana = 2,013+ 0,193 x densidad; r2 = 0,453, p = 0.0003) (Pérez-Barbería y Nores, 1994). Pirineos La conducta social del rebeco pirenaico ha sido objeto de bastantes estudios, sobre todo por parte de investigadores franceses de la escuela de Toulouse (González y Berducou, 1985; Richard-Hansen y Campan, 1992; Pepin et al., 1996b?). Se han descrito numerosos tipos de grupos atendiendo a las combinaciones de sexo-edad para su clasificación (Figura 1; Berducou y Bousses, 1985). En realidad, la única combinación grupal estable y sólida parece ser la de madre-hijo, la cual puede prolongarse hasta el segundo año de vida (Bousses y Cornaire, 1982; González y Berducou, 1985; Gerard y Richard-Hansen, 1992). También se ha descrito la formación de ciertos clanes, que corresponderían a grupos de animales que migran juntos, pero sin una gran cohesión entre ellos en el tiempo (Crampe et al., 2004).
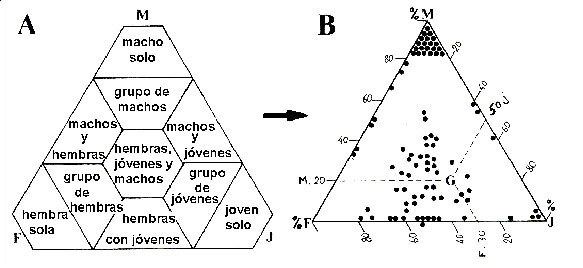
Figura 1 A. Diagrama propuesto por Berducou y Bousses (1985) para representar a los diferentes tipos de grupo de sarrio según la proporción de machos, jóvenes y hembras que entran en su composición. B. Ejemplo de distribución grupal encontrado en el Valle d’Osseau en noviembre de 1980; el punto G correspondería a un grupo matriarcal con 30% hembras, 50% jóvenes y 20% machos.
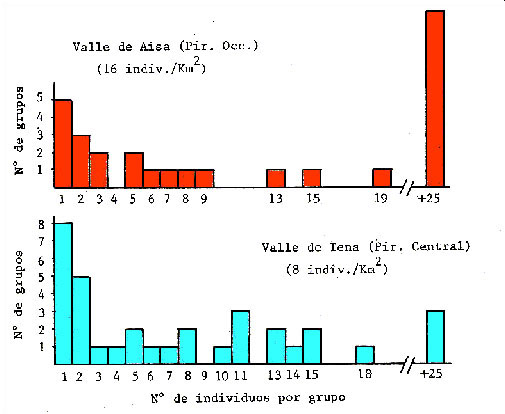
Los machos suelen estar separados de las hembras excepto en el período de celo, el cual ocurre durante el mes de noviembre. El resto del año los machos pueden ser solitarios, formar grupos de machos o grupos mixtos con las hembras. Durante el celo los machos dominantes establecen harenes de varias hembras que guardan y defienden frente a otros machos. También se han descrito tendencias al comportamiento territorial en los machos (Pepin et al., 1991; Crampe, 1997; Pérez-Barbería et al., 1997). En las grandes manadas matriarcales a veces puede observarse la formación de "guarderías" (Couturier, 1938; Krämer, 1969), que correspondería a grupos de cabritos rodeados por hembras adultas, que supuestamente ejercerían una cierta "vigilancia" sobre ellos. En invierno las hembras forman grupos que pueden incluir hembras residentes y hembras migrantes. El nivel de asociación en invierno entre hembras residentes y migrantes que utilizaban las mismas zonas en verano fue mayor que el de hembras que utilizaban zonas diferentes, lo que sugiere discriminación familiar o reconocimiento individual (Crampe et al., 2010)2. Generalmente dos son los factores importantes que se arguyen para explicar el tamaño medio de los grupos: la densidad de población (Pérez-Barbería y Nores, 1994) y el carácter abierto o cerrado del medio (Richard-Hansen et al., 1992). La elevada densidad favorece la existencia de grandes grupos matriarcales y de grupos de machos (Figura 2) (Berducou y Bousses, 1985) y el medio forestal parece favorecer la formación de grupos pequeños (Herrero et al., 1996). Sin embargo se han encontrado excepciones a este patrón general, tanto en Pirineos (Richard-Hansen et al., 1992; Crampe, 1997) como en otras subespecies (Lovari y Cosentino, 1986).
Figura 2. Comparación de la distribución del tamaño de los grupos encontrados en dos poblaciones pirenaicas de diferente densidad (según García-González et al., 1985).
La dinámica de grupos se relaciona con la densidad de población. La tasa a la que los grupos se unen aumenta al incrementarse el número de grupos. Por otro lado, la tasa a la que los grupos se dividen aumenta con su tamaño. En tercer lugar, la tasa de eventos de fusión o división aumenta cuando la densidad de población se incrementa (Pepin y Gerard, 2008). La abundancia y distribución del alimento son los parámetros que mejor explican el tamaño de los grupos (Pérez-Barbería y Nores, 1994). En invierno el alimento escaso y disperso favorece la formación de grupos pequeños, probablemente debido a que así tienden a disminuir la competencia alimentaria, como se ha sugerido para el corzo (Maublanc et al., 1987). Sin embargo, desde la primavera hasta el otoño la disponibilidad de alimento es mayor y los grupos crecen en tamaño. Las grandes agregaciones del verano pueden estar asociadas con la distribución restringida de las zonas de alimentación preferente, más que a complejas interacciones sociales. Estas zonas suelen ser pequeños valles colgados, con pastos de alta calidad (del tipo Primulion), y alejados del tránsito humano y de las zonas intensamente utilizadas por el ganado (García González et al., 1985).
Referencias Bousses, P., Cornaire, S. (1982). L'ontogenese du comportement de jeune isard. Documents scientifiques du Parc National des Pyrénées, 10: 1-122. Couturier, M. A. J. (1938). Le chamois. B. Arthaud, Grenoble. Crampe, J. P. (1997). Caractéristiques bio-démographiques d'une population d'isards (Rupicapra p. pyrenaica) non-chasée dans le Parc National des Pyrénées, en vallée de Cauterets. Documents scientifiques du Parc National des Pyrénées, 31: 1-169. Crampe, J. P., Bon, R., Gerard, J. F., Gonzalez, G. (2010). Do isard females associate in winter on the basis of summer bonds? Galemys, 22: 531-543. Crampe, J. P., Bon, R., Gerard, J. F., Serrano, E., Caens, P., Florence, E., González, G. (2007). Site fidelity, migratory behaviour, and spatial organization of female isards (Rupicapra pyrenaica) in the Pyrenees National Park, France. Canadian Journal of Zoology, 85 (1): 16-25. Crampe, J. P., Caens, P., Florence, E., Gérard, J. F., González, G., Serrano, E. (2004). Résultats préliminaires sur l'organisation spatiale de femelles d'isard (Rupicapra pyrenaica) dans une population du Parc National des Pyrénées. En: Herrero, J., Escudero, E., Luco, D.F.d., García-González., R. (Eds.). El sarrio: biología, patología y gestión. Consejo Protección de la Naturaleza de Aragón, Gobierno de Aragón, Zaragoza. Elsner-Schack, Von, I. (1985). Seasonal changes in the size of chamois groups in the Ammergauer mountains, Bavaria. Pp. 148-153. En: Lovari, S. (Ed.). The Biology and management of mountain ungulates. Croom-Helm, London. García-González, R., Herrero, J., Hidalgo, R. (1985). Estimación puntual de diversos parámetros poblacionales y distributivos del sarrio en el Pirineo Occidental. Pirineos, 35: 53-63. Gerard, J.F., Richard-Hansen, C. (1992). Social affinities as the basis of the social organization of a Pyrenean chamois (Rupicapra pyrenaica) population in an open mountain range. Behavioural Processes, 28: 111-122. Gonzalez, G., Berducou, C. (1985). Les groupes sociaux d' isards et de mouflons au massif du Carlit (Pyrénées Orientales). Gibier Faune Sauvage, 4: 85-102. Herrero, J., Garin, I., García-Serrano, A., García-González, R. (1996). Habitat use in a Rupicapra pyrenaica pyrenaica forest population. Forest Ecology and Management, 88: 25-30. Hirth, D. H. (1977). Social behaviour of white-tailed deer in relation to habitat. Wildlife Monographs, 53: 1-55. Jordá, P. J. (1981). Parámetros de gregarismo del gamo (Dama dama) en el coto de Doñana. Doñana, Acta Vertebrata, 8: 237-289. Krämer, A. (1969). Organisation sociale et comportement social d'une population de chamois (Rupicapra rupicapra L.) des Alpes. Z. Tierpsychology, 26: 889-964. Lagory, K. E. (1986). Habitat, group size, and the behaviour of white-tailed deer. Behaviour, 98: 168-179. Levet, M., Appolinaire, J., Catusse, M., Thion, N. (1995). Demographic data, spacial behaviour and dispersion of an Isard (Rupicapra pyrenaica pyrenaica) population in stage of colonization. Mammalia, 59: 489-500. Loison, A., Jullien, J. M., Menaut, P. (1999). Subpopulation structure and dispersal in two populations of chamois. Journal of Mammalogy, 80: 620-632. Lovari, S., Cosentino, R. (1986). Seasonal habitat selection and group size of the Abruzzo chamois (Rupicapra pyrenaica ornata). Bool. Zool., 53: 73-78. Maublanc, M.-L., Bideau, E., Vincent, J.-P. (1987). Flexibilité de l'organisation sociale du Chevreuil en fonction des caractéristiques de l'environnement. Rev. Ecol. (Terre Vie), 42: 109-133. Pepin, D., Gerard, J. F. (2008). Group dynamics and local population density dependence of group size in the Pyrenean chamois, Rupicapra pyrenaica. Animal Behaviour, 75 (2): 361-369. Pepin, D., González, G., Bon, R. (1991). Le Chamois et L'isard. Rev. Ecol. (Terre Vie), Suppl., 6: 111-153. Pepin, D., Lamerenx, F., Chadelaud, H. (1996b). Diurnal grouping and activity patterns of the Pyrenean chamois in winter. Ethology, Ecology & Evolution, 8: 135-145. Pérez-Barbería, F. J., Oliván, M., Osoro, K., Nores, C. (1997). Sex, seasonal and spatial differences in the diet of Cantabrian chamois Rupicapra pyrenaica parva. Acta Theriologica, 42: 37-46. Pérez-Barbería, F. J., Nores, C. (1994). Seasonal variation in group size of Cantabrian chamois in relation to escape terrain and food. Acta Theriologica, 39: 295-305. Richard-Hansen, C., Campan, R. (1992). Social environment of Isard Kids, Rupicapra p. pyrenaica, during their ontogeny. Z. Säugetierkunde, 57: 351-363. Richard-Hansen, C., González, G., Gerard, J. F. (1992). Structure sociale de l'isard (Rupicapra pyrenaica) dans trois sites pyrénéens. Gibier Faune Sauvage, 9: 137-149. Solá, J. (2004). El sarrio en el Principado de Andorra. Estado de las poblaciones, gestión y perspectivas. Segundas Jornadas técnicas sobre el sarrio en Aragón. Gobierno de Aragón, Jaca, 31 de marzo al 2 de abril 2004.
Francisco Javier Pérez-Barbería Ricardo García-González Borja Palacios Alberti Fecha de publicación: 16-06-2004 Revisiones: 24-07-2008; 28-10-2010 Otras contribuciones: 2. Alfredo Salvador. 10-08-2017 Pérez-Barbería, F. J., García-González, R., Palacios, B. (2017). Rebeco – Rupicapra pyrenaica. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Barja, I. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|
|
|
|
|
|