|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Keywords: Bearded Vulture, diet, food preferences, ossuaries.
Ecología trófica Composición general de la dieta Su dieta se compone de huesos y carne de cadáveres de vertebrados, fundamentalmente mamíferos, pero también aves y reptiles. Ocasionalmente captura pequeños vertebrados (Glutz von Blotzheim et al., 1971; Hiraldo et al., 1979; Cramp y Simmons, 1980). Hace 10.000-14.000 años la dieta en España estaba compuesta de Rupicapra pyrenaica, Capra pyrenaica, Cervus elaphus y Capreolus capreolus. Por el contrario, en tiempos históricos (1940-1950) y en la actualidad (2008) predominan cabras y ovejas (Margalida y Martín-Arroyo, 2013)1. La dieta en España, según una recopilación preliminar, está basada en mamíferos domésticos y silvestres (Capra pyrenaica, Rupicapra pyrenaica, Vulpes vulpes, Rattus sp., Oryctolagus cuniculus), aves (Columba palumbus, Streptopelia turtur, Lagopus mutus, Alectoris rufa, Upupa epops, Pyrrhocorax pyrrhocorax, Pica pica, Sylvia atricapilla, Phylloscopus collybita) y reptiles (Timon lepidus) (Hiraldo et al., 1979). Según otro análisis, de un total de 152 restos recogidos en nidos y rompederos pirenaicos, el 88% son mamíferos, el 7% aves y el 0,7% reptiles. Entre los mamíferos predominan los ungulados domésticos y el rebeco como ungulado salvaje más representado en la dieta. Entre las aves destaca la paloma torcaz (Heredia, 1991d). Un estudio más detallado, basado en observaciones con telescopio y videocámaras de 677 presas aportadas a 8 nidos, indica que la dieta durante el periodo reproductor se basa en mamíferos (93%), sobre todo ungulados de tamaño medio (principalmente Ovis/Capra y Rupicapra pyrenaica), representando las aves el 6% y los reptiles el 1% (Margalida et al., 2009). Respecto a la captura de presas vivas, se ha registrado en España la captura de un pollo de gaviota (Larus argentatus) y de una culebra (Hiraldo et al., 1979). Se conoce el consumo de las propias egagrópilas por los adultos (Margalida y Bertran, 1996) y se ha citado el consumo ocasional de egagrópilas de búho real (Heredia et al., 1990). La mayoría de los estudios de la dieta en España han sido realizados en Pirineos y son escasos los datos disponibles de otras áreas donde la especie se extinguió, como la sierra de Cazorla (Cano y Valverde, 1959; Otero et al., 1979). González y González (1984) encontraron en un rompedero de Cazorla restos de muflón, Cervidae indet., Capra sp. y Caprinae indet.. La comparación de restos en nidos del sur de España durante el periodo 1940-1950 con los examinados en Pirineos durante 2000-2001 muestra una dieta similar, basada en mamíferos domésticos de tamaño medio (oveja y cabra) (Margalida et al., 2009). A pesar de que la dieta es fundamentalmente ósea, existen diferencias entre la alimentación de los adultos y la seleccionada por éstos durante la etapa de crianza para alimentar al pollo en la que las presas cárnicas a través de pequeños cadáveres, adquiere mayor relevancia (Margalida y Bertran, 1997, Margalida & Bertran, 2001, Margalida et al., 2001). Disponibilidad de alimento y dieta Se ha señalado que una de las causas de regresión de poblaciones de aves carroñeras ibéricas es, además de la alteración del hábitat, la baja disponibilidad de alimento (Donázar, 1993). Se ha estimado que la biomasa anual proporcionada por los cadáveres de ungulados domésticos y salvajes supera los requerimientos energéticos estimados para las parejas reproductoras, tanto durante el periodo reproductor como durante el resto del año (Canut et al., 1987; LorenteVillanueva, 1996; Margalida et al., 1997). La mayor cantidad y diversidad de recursos tróficos se dan en la zona axial o alto Pirineo. En las sierras exteriores hay una fuerte dependencia de los muladares y rebaños de ganado doméstico (Heredia, 1991d). Sesgos en los métodos usados para la determinación de la dietaEl mayor problema relacionado con el estudio de la dieta del quebrantahuesos es que se alimenta de los restos de animales muertos, especialmente huesos, los cuales son difíciles de identificar y pueden ser digeridos por completo; las egagrópilas en el quebrantahuesos no son abundantes y ocasionalmente son recicladas para aprovechar los restos óseos no digeridos en primer término (Margalida y Bertran 1996) y la especie utiliza despensas y rompederos donde prepara el alimento, generalmente los restos óseos de mayor tamaño pertenecientes a grandes ungulados, dispersando así parte de la información (Heredia 1991d, Bertran y Margalida 1996, Margalida y Bertran 2001), A este respecto, los sesgos o diferencias en la determinación de la dieta utilizando diferentes metodologías se han podido estimar testando las diferencias existentes entre métodos de observación directa como son las videocámaras y los telescopios (Margalida et al., 2005) así como los sesgos inherentes a las diferentes metodologías que pueden permitir determinar el espectro trófico de la especie: los restos aportados al nido, los restos presentes en el nido, los restos recogidos tras la crianza y los restos recogidos en rompederos (Margalida et al. 2007). En el estudio que compara la dieta de la especie con métodos de observación directa (telescopios vs vídeo-cámaras) se monitorizaron intensivamente cuatro parejas focales en las que se aplicaron tres metodologías distintas: la observación de presas aportadas al nido, la observación de restos presentes en el nido (que no contabiliza los aportados durante la jornada de observación) y la recogida de restos tras la crianza. Un cuarto método que consistió en sumar las presas aportadas con las presentes en el nido, también fue testado puesto que a priori permite aumentar el tamaño de muestra sin provocar sesgos (Margalida et al., 2005). Para determinar los sesgos se compararon las frecuencias obtenidas en las presas presentes en el nido (restos que eran observados en el interior del nido durante el seguimiento de la pareja) y las recogidas tras la crianza (restos recogidos en el nido y a pie de nido tras finalizar la crianza) con las observadas aportadas al nido (como método más objetivo en la determinación de la dieta). Los restos recogidos sobrevaloraron las presas de gran tamaño pertenecientes a los géneros Bos/Equus así como la proporción de aves, infravalorando los ungulados de tamaño medio de los géneros Ovis/Capra. Por otro lado, las presas presentes en el nido infravaloraron los micromamíferos. Cuando combinamos restos presentes en el nido con restos aportados, los datos obtenidos con respecto a las presas aportadas mostraron la ausencia de diferencias. Estos resultados sugieren que el método más objetivo para la determinación de la dieta es la observación directa y que éste puede ser optimizado si lo combinamos con los restos presentes en el nido, ya que su inclusión no provoca sesgos y aumenta el tamaño muestral (Margalida et al., 2005). Un posterior estudio incrementando el número de parejas y considerando también los restos recogidos en rompederos mostró diferencias significativas tanto a nivel anatómico como taxonómico (Margalida et al. 2007, Tabla 1). Así, con respecto a las presas aportadas, los restos presentes en el nido infravaloraron los mamíferos pequeños y micromamíferos mientras que los restos recogidos tras la crianza infravaloraron los mamíferos de tamaño medio y los micromamíferos, sobrevalorando los restos de mamíferos grandes y las aves. Finalmente, los rompederos fueron la metodología menos objetiva, difiriendo significativamente en la práctica totalidad de las categorías consideradas. Tabla 1. Proporción de tipos de presa en 8 parejas de quebrantahuesos de Pirineos durante 1995-2004 según cuatro tipos de métodos: presas aportadas durante el periodo de observación (n= 526), presas presentes en el nido al comienzo del periodo de o bservación (n= 494), restos encontrados en el nido al finalizar la reproducción (n= 263) y restos encontrados en rompederos (n= 252). Mamíferos de tamaño medio incluye oveja, cabra, sarrio y ciervo; mamíferos de tamaño grande incluye vaca y caballo; mamíferos pequeños incluye conejo, liebre y zorro. Según Margalida et al. (2007).
Dieta y selección de alimentoLa información sobre la alimentación del quebrantahuesos en los Pirineos proviene de observaciones esporádicas (Terrasse et al., 1961; Suetens, 1974; Terrasse y Terrasse, 1974; Hiraldo et al., 1979; Terrasse, 1981; Heredia y Herrero, 1992), de restos recogidos en los rompederos (Heredia, 1974; Boudoint, 1976) y de contenidos estomacales (Palaus, 1959). La alimentación de la especie durante la crianza no es muy conocida (Suetens y Van Groenendael, 1973). Sin embargo, estudios recientes (Margalida et al., 2005) realizados en Pirineos arrojan de forma preliminar luz sobre este aspecto. Como hemos descrito anteriormente, la observación directa es el método más objetivo para cuantificar la dieta de ésta y otras especies. Como refleja la Tabla 2, la dieta de la especie se fundamenta en los mamíferos (95.2%). Las aves sólo representan el 4% de los restos identificados y los reptiles el 0.8% restante. Entre los mamíferos, los restos de ungulados domésticos y salvajes (67% de las presas identificadas) suponen el grueso de la dieta, si bien como especie oportunista, existen diferencias entre territorios y la especie puede presentar un espectro trófico muy variado adaptado a la disponibilidad de recursos presentes en cada territorio.
Tabla 2. Dieta del quebrantahuesos en los Pirineos durante la crianza (n = 8 territorios). Datos obtenidos mediante observación directa con videocámaras y telescopios (Margalida et al., 2005).
Para conocer la selección de alimento con respecto a la disponibilidad de presas presentes en el territorio es necesario realizar estimas de los recursos disponibles contrastados con los seleccionados por la especie. El primer estudio que ha contrastado la disponibilidad trófica teórica con la seleccionada aportada al nido se realizó en una pareja pirenaica (Margalida y Bertran, 1997). Mediante observación directa con telescopios se comparó la dieta de la pareja con respecto a la disponibilidad teórica de carroña presente en el área de campeo habitual explotada por los adultos (Tabla 3).
Tabla 3. Presencia, abundancia y disponibilidad de los recursos potencialmente utilizables por una pareja de quebrantahuesos durante la crianza en los Pirineos (Margalida y Bertran, 1997).
Tabla 4. Relación de las presas aportadas al nido por una pareja de quebrantahuesos durante la crianza (Margalida y Bertran 1997).
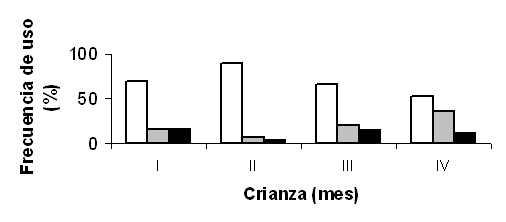
Los 59 aportes correspondientes a especies domésticas constituyeron la mayor parte de la dieta del pollo (el 92,2%). Los aportes más consumidos fueron los restos de oveja/cabra (59,4%), con 32 huesos (20 extremidades, 7 columnas, 1 cráneo y 4 huesos) y seis pieles, seguidas de los de conejos Oryctolagus cuniculus var. dom. (25%), con 16 restos. El resto de presas domésticas identificadas fueron: un cráneo de perro Canis sylvestris, una pata de ternero, un hueso de vacuno/equino y una escápula y un hueso de porcino. Las cuatro presas no domésticas fueron: el cadáver seco de un zorro Vulpes vulpes, un mamífero no identificado, una chocha perdiz Scolopax rusticola, una paloma no identificada y un ofidio no identificado (Tabla 4), que en conjunto representaron únicamente el 8% del total de presas identificadas. Los restos óseos de porcino y de vacuno/equino aparecieron con frecuencia muy baja (3% cada uno). El análisis de la selección de presas demostró la existencia de una tendencia positiva respecto los grupos ovino/caprino y cunícola, y una tendencia negativa en la selección del grupo formado por animales de talla más grande (vacuno/equino y, en menor medida, porcino). De los 75 aportes registrados al nido, el 50,7% fueron restos óseos, un porcentaje significativamente inferior respecto a los datos obtenidos por otros autores que habían aplicado la misma metodología (Suetens y van Groenendael 1973), lo que sugiere que las presas con contenido cárnico pueden tener más importancia en la dieta del pollo que en la de los adultos. Los resultados muestran que, tal y como sugieren otros autores (Suetens y van Groenendael, 1973; Otero et al., 1979; Thibault et al., 1993), los animales domésticos son una parte muy importante de la dieta del quebrantahuesos. A pesar de ello, probablemente hay diferencias entre parejas y subpoblaciones. La presencia de animales domésticos es probablemente inferior en las parejas pirenaicas, que harán más uso de los ungulados salvajes como el rebeco (Heredia 1991d, Donázar, 1993; García y Margalida, 1998), y donde la menor previsibilidad de alimento posiblemente acentuará el carácter oportunista de la especie (Hiraldo et al., 1979). Un estudio reciente, basado en observaciones con telescopio y videocámaras de las presas aportadas a 8 nidos, indica que los restos de grandes presas (vacas y caballos) son seleccionados negativamente y que los quebrantahuesos prefieren comer las patas en comparación con otras partes del cuerpo (Margalida et al., 2009). Uso y funcionalidad de los rompederosLos ejemplares adultos pueden ingerir huesos de un tamaño de 250 mm de largo y 35 mm de ancho sin dificultad aparente. El problema que supone la ingestión de huesos mayores lo ha resuelto con la utilización de los llamados rompederos. Los rompederos son superficies rocosas que son utilizados por los quebrantahuesos para la rotura y preparación de los restos óseos. Aunque algunos rompederos sólo se utilicen ocasionalmente, la especie dispone, en las inmediaciones de los nidos, de ubicaciones concretas donde regularmente lleva a cabo esta actividad. Los rompederos acostumbran a estar ubicados en zonas expuestas al viento y con pendiente. Estas características facilitan, por un lado, que el ave pueda remontar fácilmente durante las sesiones de rotura, minimizando el desgaste energético que requiere la realización de esta actividad y, por otro, que se incrementen las posibilidades de fragmentación como consecuencia del rebote de los restos óseos (Heredia, 1991d). Los rompederos están situados a altitudes entre 860 y 2.300 m (Hiraldo et al., 1979). Los rompederos son utilizados durante todo el año, pero es durante la crianza cuando se acentúa su importancia (Heredia, 1991d; Margalida y Bertran, 2001), dado que estas ubicaciones se usan con más regularidad para preparar el alimento para el pollo. Los quebrantahuesos utilizan un término medio de 2 ± 0,58 rompederos (rango 1-3) ubicados a una distancia media de los nidos de 789,3 ± 376,5 m (rango= 175-2400 m; n = 14) (Margalida y Bertran, 2001). Los rompederos están situados a altitudes entre 860 y 2.300 m. La altura de suelta de los huesos varía entre 20 y 150 m (Hiraldo et al., 1979). El 84,9% de los restos encontrados en osarios pertenecen a ungulados de mediado tamaño, el 11,6% a mamíferos grandes, el 2,5% a carnívoros, y el 1% a Suidae. Los restos encontrados son sobre todo escápulas (14,9%), vértebras (13,2%), cráneos (12,5%), tibias (10,7%, mandíbulas (9,3%), costillas (8,3%) y húmeros (7,2%). Muchos de estos restos permanecen en los osarios sin ser consumidos, caracterizándose por el predominio de partes esqueléticas poco nutritivas (Margalida, 2008b). La comparación de los huesos aportados al nido (seleccionados), con los huesos disponibles en comederos y los observados en osarios sugiere que prefieren aquellos huesos con mayor contenido en ácido oleico independientemente de su longitud, aunque la morfología del hueso y su influencia en su manejo e ingestión puede jugar un papel secundario (Margalida, 2008a). El tiempo medio empleado para romper los huesos fue de 5,3 ± 4,2 minutos (rango= 1-17; n = 50 sesiones de rotura de huesos) y durante las sesiones de rotura, el número medio de lanzamientos realizados para conseguir la fragmentación o la desarticulación del resto fue de 4,5 ± 5,8 (rango= 1-35; n = 86 sesiones de rotura de huesos). De las 94 visitas registradas en los rompederos, en el 71,3% de los casos la utilización del rompedero estuvo asociada a la alimentación del pollo, en el 18,1% a la alimentación del adulto y en el 10,6% al almacenamiento de alimento. Durante la crianza, los rompederos son utilizados principalmente para adecuar el tamaño del alimento a las necesidades y a las posibilidades de ingestión del pollo y, en menor medida, para que el adulto se prepare su propia comida. La preparación del alimento para el pollo mostró diferencias significativas en la frecuencia temporal de utilización de los rompederos, siendo significativamente inferior durante el primer mes de vida del pollo (0,023 ± 0,059 sesiones hr-1) respecto al segundo y al tercer mes (0,072 ± 0,123 y0,073 ± 0,108, respectivamente), pero similar al cuarto (0,027 ± 0,052) (Figura 1).
Figura 1. Variación temporal en el uso y funcionalidad de los rompederos para la preparación del alimento. Columnas blancas: preparación del alimento para el pollo; columnas grises: preparación del alimento para el adulto; columnas negras: almacenamiento del alimento (Margalida y Bertran, 2001).
La variación temporal del uso de los rompederos podría ser debida a una posible variación cualitativa de la dieta del pollo. Aunque en cautividad se ha comprobado que el pollo pocas horas después de la eclosión puede ingerir pequeños restos óseos combinados con carne, durante el primer mes las presas cárnicas probablemente pueden adquirir especial relevancia en la alimentación. El tiempo medio de estancia en los rompederos dedicado a la preparación del alimento no difirió entre ambos periodos (< 60 días: 4,5 ± 3,8 minutos, n = 27; > 61 días, 6 ± 4,5 minutos, n = 23). La significativa disminución de la frecuencia de visitas a los rompederos y del tiempo dedicado a cebar el pollo durante las dos primeras semanas (Margalida y Bertran, 2001) también sugieren la presencia de presas cárnicas en la dieta. La progresión en la capacidad de tragar por parte del pollo se traduciría en un aporte mayor de restos óseos durante el segundo y el tercer mes. Esta variación en la dieta justificaría un incremento del uso de los rompederos para la preparación de alimento. El hecho de que decrezca bruscamente el uso de rompederos durante el mes precedente al abandono del nido puede estar asociado a dos factores. Por un lado, está la capacidad de ingerir huesos por parte del pollo, que es considerable, y permite a los adultos disminuir el tiempo de preparación en los rompederos. Por otro lado, durante esta etapa se produce la trashumancia, periodo en el que la disponibilidad de recursos tróficos es cuantitativamente la más abundante (Margalida et al., 1997). La abundancia relativa de alimento y la consecuente previsibilidad espacial y temporal de su búsqueda permiten a la especie seleccionar la tipología de restos más adecuados, lo que reduciría el tiempo de preparación en los rompederos. La hipótesis de la variación temporal de la calidad del alimento también estaría corroborada por las diferencias observadas en el comportamiento de los adultos respecto al uso de los rompederos. Mientras que durante los dos primeros meses el alimento es preparado con mayor frecuencia antes de ser aportado al nido, durante los dos últimos meses es más habitual ir al rompedero después de haber aportado los restos al nido. Estas diferencias se deberían al hecho de que, durante la primera mitad de la crianza, las limitaciones del pollo en la ingestión de los huesos obligarían al adulto a preparar la comida antes de entrar en el nido. Los restos cárnicos y los pequeños fragmentos óseos son consumidos por el pollo pero generalmente es el adulto el que acaba ingiriendo el hueso (obs. pers.). Almacenaje del alimentoEl quebrantahuesos puede almacenar alimento para comerlo días o semanas más tarde. El hecho de que los huesos se mantengan en buenas condiciones para el consumo durante largos periodos hace que la especie pueda utilizar sitios donde almacenarlos. Los posaderos donde habitualmente prepara e ingiere los restos, los rompederos y los nidos son las ubicaciones más utilizadas para guardar el alimento. Cuando las condiciones meteorológicas no son favorables o cuando los recursos escasean, el quebrantahuesos visita los rompederos para abastecerse de los restos almacenados. Esto incrementa las interacciones intraespecíficas (y ocasionalmente interespecíficas, véase Bertran y Margalida, 1997; Margalida y Bertran, 2003), dado que estos puntos previsibles en el espacio son visitados frecuentemente por otros quebrantahuesos, principalmente por individuos de la población flotante (Bertran y Margalida, 1996). El almacenaje de la comida se lleva a cabo en rompederos cercanos al nido, lo que permitiría a los adultos reproductores un control regular que evitaría o reduciría el riesgo del pirateo por parte de conespecíficos (Bertran y Margalida, 1996; Margalida y Bertran, 2003), en nidos no utilizados y en cuevas (Heredia, 1991d). Cleptoparasitismo El quebrantahuesos hostiga a otras aves hasta conseguir que suelten la presa, apoderándose de ella. Se ha visto al quebrantahuesos acosando a águilas reales (Aquila chrysaetos), alimoches (Neophron percnopterus), cuervos (Corvus corax) y chovas (Phyrrocorax sp.) (Hiraldo et al., 1979, Margalida y Bertran, 2003). La mayoría de los casos de cleptoparasitismo se refieren a inmaduros, que a menudo roban restos de adultos en osarios y nidos, más raramente en vuelo (Margalida y Bertran, 2003).
Referencias Bertran, J., Margalida, A. (1996). Patrón anual de observaciones de quebrantahuesos (Gypaetus barbatus) de diferentes grupos de edad en los sectores de nidificación. Alauda, 64: 171-178. Bertran, J., Margalida, A. (1997). Griffon vultures (Gyps fulvus) ingesting bones in the ossuaries of Bearded vultures (Gypaetus barbatus). J. Raptor Res., 31: 287-288. Boudoint, Y. (1976). Techniques de vol el cassage d'os chez le gypaète barbu Gypaetus barbatus. Alauda, 44: 1-21. Cano, A., Valverde, J. A. (1959). Notas sobre el quebrantahuesos Gypaetus barbatus en la sierra de Cazorla. Ardeola, 5: 121-126. Canut, J., García, D., Heredia, R., Marco, J. (1987). Status, características ecológicas, recursos alimenticios y evolución del quebrantahuesos (Gypaetus barbatus) en la vertiente sur de los Pirineos. Acta Biologica Montana, 7: 83-99. Cramp, S., Simmons, K. E. L. (Eds.) (1980). Handbook of the Birds of Europe the Middle East and North Africa. The Birds of the Western Palearctic. Volume II. Hawks to Bustards. Oxford University Press, Oxford. Donázar, J. A. (1993). Los buitres ibéricos. Biología y conservación. J. M. Reyero, Madrid. García, D., Margalida, A. (1998). Avaluació de la disponibilitat tròfica per al Trencalòs (Gypaetus barbatus) al Parc Nacional d'Aigüestortes i Estany de Sant Maurici: importància de les poblacions d'Isards (Rupicapra pyrenaica). Pp. 179-188. En: IV Jornades sobre Recerca al Parc Nacional d'Aigüestortes: Generalitat de Catalunya, Departament de Medi Ambient, Lleida. Glutz von Blotzheim, U. N., Bauer, K. M., Bezzel, E. (1971). Handbuch der Vögel Mitteleuropas. Band 4. Falconiformes. Akademische Verlagsgessellschaft, Wiesbaden. González, L. M., González, J. L. (1984). Sobre la distribución pasada y alimentación del quebrantahuesos (Gypaetus barbatus L. 1758) en España. Doñana Acta Vertebrata, 11 (1): 141-143. Heredia, R. (1974). Nota sobre la alimentación del quebrantahuesos (Gypaetus barbatus). Ardeola, 19: 345-346. Heredia, R. (1991d). Alimentación y recursos alimenticios. Pp. 79-89. En: Heredia, R., Heredia, B. (Eds.). El quebrantahuesos (Gypaetus barbatus) en los Pirineos: características ecológicas y biología de la conservación. ICONA, Madrid. Heredia, R., Donázar, J. A., Ceballos, O. (1990). Ingestion of eagle owl Bubo bubo pellets by lammergeiers Gypaetus barbatus. Ibis, 132 (1): 127. Heredia, R., Herrero, J. (1992). Bearded vulture (Gypaetus barbatus) and Alpine marmot (Marmota marmota) interactions in southern Pyrenees. Pp. 227-229. En: Bassano, B., Durio, P., Gallo Ursi, U., Macchi, E. (Eds.).First Int. Symp. Alpine Marmot. Hiraldo, F., Delibes, M., Calderón, J. (1979). El quebrantahuesos Gypaetus barbatus(L.). Sistemática, Taxonomía, Biología, Distribución y Protección. Monografías, 22. Instituto para la Conservación de la Naturaleza, Madrid. Lorente Villanueva, L. (1996). Disponibilidad de recursos alimenticios para el quebrantahuesos (Gypaetus barbatus) en el Pirineo aragonés. Lucas Mallada, 8: 109-119. Margalida, A. (2008a). Bearded vultures (Gypaetus barbatus) prefer fatty bones. Behavioral Ecology and Sociobiology, 63 (2): 187-193. Margalida, A. (2008b). Presence of bone remains in the ossuaries of bearded vultures (Gypaetus barbatus): storage or nutritive rejection? Auk, 125 (3): 560-564. Margalida, A., Bertran, J. (1996). Quebrantahuesos (Gypaetus barbatus) ingiriendo sus propias egagrópilas. Butlletí del Grup Catalá d’Anellament, 13: 49-51. Margalida, A., Bertran, J. (1997). Dieta y selección de alimento de una pareja de quebrantahuesos (Gypaetus barbatus) en los Pirineos durante la crianza. Ardeola, 44 (2): 191-197. Margalida, A., Bertran, J. (2001). Function and temporal variation in use of ossuaries by bearded vultures (Gypaetus barbatus) during the nestling period. Auk, 118 (3): 785-789. Margalida, A., Bertran, J. (2003). Interspecific and intraspecific kleptoparasitic interactions of the bearded vulture (Gypaetus barbatus) at nesting areas. Journal of Raptor Research, 37 (2): 157-160. Margalida, A., Bertran, J., Boudet, J. (2005). Assessing the diet of nestling bearded vultures: a comparison between direct observation methods. Journal of Field Ornithology, 76 (1): 40-45. Margalida, A., Bertran, J., Heredia, R. (2009). Diet and food preferences of the endangered bearded vulture Gypaetus barbatus: a basis for their conservation. Margalida, A., Bertran, J., Heredia, R., Boudet, J., Pelayo, R. (2001). Preliminary results of the diet of Bearded Vultures (Gypaetus barbatus) during the nestling period and applications in conservation and management measures. Pp. 59-62. En: Sakoulis, A., Probonas, M., Xirouchakis, S. (Eds.). Proceedings of the 4th Workshop of Bearded Vulture. Natural History Museum of Crete, Irakleio. Margalida, A., Boudet, R., Heredia, R., Bertran, J. (2002a). Videocámaras para la monitorización de la nidificación del Quebrantahuesos (Gypaetus barbatus). Ecología, 16: 325-333. Margalida, A., García, D., Heredia, R. (1997). Estimación de la disponibilidad trófica para el quebrantahuesos (Gypaetus barbatus) en Cataluña (NE España) e implicaciones sobre su conservación. Doñana Acta Vertebrata, 24 (1-2): 235-243. Margalida, A., Heredia, R., Boudet, R. (2002b). Caméras vidéo pour l’etude de la biologie de la reproduction du Gypaète barbu (Gypaetus barbatus): un premier bilan. Pp. 115-121. En: LPO/FIR (Eds.). Proceedings of the International Conference “Conservation of Bearded Vulture populations”. LPO, Paris. Margalida, A., Mañosa, S., Bertran, J., García, D. (2007). Biases in studying the diet of the bearded vulture. Journal of Wildlife Management, 71 (5): 1621-1625. Margalida, A., Marín-Arroyo, A. B. (2013). Dietary habits in the endangered Bearded Vulture Gypaetus barbatus from Upper Pleistocene to modern times in Spain: a paleobiological conservation perspective. Bird Conservation International, 23 (4): 469-476. Margalida, A., Sánchez-Zapata, J. A., Eguia, S., Marin Arroyo, A. B., Hernández, F. J., Bautista, J. (2009). Assessing the diet of breeding bearded vultures (Gypaetus barbatus) in mid-20th century in Spain: a comparison to recent data and implications for conservation. European Journal of Wildlife Research, 55 (4): 443-447. Otero, C., Castién, R., Senosiain, A., Portillo, F. (1978). Fauna de Cazorla. Vertebrados. Monografías, 19. Instituto para la Conservación de la Naturaleza, Madrid. Palaus, F. J. (1959). Notas para el estudio del Gypaëtus barbatus en los Pirineos Orientales. Misc. Zool., 1: 97-100. Suetens, W. (1974). Otra breve nota sobre alimentación de Gypaetus barbatus. Ardeola, 19: 347. Suetens, W., Groenendael, P. V. (1973). Bijdrage tot ecologi en de ethologie van de lammergeier, Gypaetus barbatus aureus (Hablizl). Gerfaut, 62: 203-214. Terrasse, J. F. (1981). Recensement, succès de reproduction et alimentation du gypaète (Gypaetus barbatus) dans les Pyrénées françaises. Pp. 76-78. En : Cheylan, G., Thibault, J. C. (Eds.). Rapaces méditerranéens, 1. Aix-en-Provence. Terrasse, J. F., Terrasse, M. (1974). Comportement de quelques rapaces nécrophages dans les Pyrénées. Nos Oiseaux, 356: 289-299. Terrasse, J. F., Terrasse, M., Boudoint, Y. (1961). Observations sur la reproduction du voutour fauve, du percnoptère et du gypaète barbu dans les Basses-Pyrénées. Alauda, 29: 1-24. Thibault, J. C., Vigne, J. D., Torre, J. (1993). The diet of young Lammergeiers Gypaetus barbatus in Corsica: its depedence on extensive grazing. Ibis, 135: 42-48. Antoni Margalida Fecha de publicación: 11-02-2011 Otras contribuciones: 1. Alfredo Salvador. 11-10-2016 Margalida, A. (2016). Quebrantahuesos – Gypaetus barbatus. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Morales, M. B. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||