|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Balearic Shearwater, reproduction, demography.
Biología de la reproducción La pardela balear frecuenta las colonias de cría ya desde agosto, aunque el grueso de la población reproductora retorna en febrero y marzo, cuando las hembras llevan a cabo la puesta. La Pardela Balear pone un solo huevo y no se conocen puestas de reposición. Los dos individuos de la pareja se alternan en la incubación del huevo durante 48-52 días, extendiéndose el período de cuidado del pollo hasta los 60-70 días. El vuelo de los pollos se produce hacia finales de junio (Oro et al., 2004; Ruiz y Martí, 2004). Se conoce desde hace poco la sorprendente existencia de tríos de cría, aunque los detalles son todavía una incógnita (Genovart et al., 2008). En las Islas Baleares el tamaño de la mayoría de los huevos está entre 53 y 59 cm3 (media= 56.9; DS = 5.2; N=209). El tamaño del huevo varió entre tres poblaciones locales vecinas y no con los años, resultado inesperado a una escala espacial tan pequeña, ya que las colonias de estudio presentaban características ecológicas similares (Louzao et al., 2008; Tavecchia et al., 2008). La variabilidad espacial en el tamaño del huevo podría explicarse parcialmente por diferencias interpoblacionales en el nivel trófico en el que se alimentan los adultos reproductores (valores promedio del isótopo d15N). Tanto las diferencias interpoblacionales en el nivel trófico como en el tamaño del huevo mostraron el mismo patrón general, es decir, los huevos más grandes estarían relacionados con valores promedio de d15N mayores, sugiriendo una conexión nutricional entre ambos. Al mismo tiempo los dos factores, el nivel trófico y el tamaño del huevo podrían estar influenciados por el tamaño corporal parental (potencial indicador de un componente genético) puesto que éste podría mediar en la selección de las presas y condicionar el tamaño del huevo (Louzao et al., 2008). La productividad (entendida como porcentaje de pollos volados respecto al número de parejas reproductoras) se sitúa entorno al 60% (Louzao et al., 2006a). No se dispone de datos relativos a la supervivencia de los individuos jóvenes e inmaduros. La productividad (entendida como el éxito de eclosión y reproductor) varió con los años y de manera similar en todas las colonias. Así, la disponibilidad de alimento, tanto de pequeños peces pelágicos como de descartes de la pesca de arrastre en la zona del delta del Ebro (principal zona de alimentación de la pardela balear), también influyó positivamente en la productividad. La variabilidad temporal de la productividad fue probablemente debida a los cambios anuales en la disponibilidad de pequeños peces pelágicos, que exhiben estocasticidad temporal, y descartes pesqueros (Louzao et al., 2006b). Algunos datos históricos sobre el éxito de eclosión y reproductor de la pardela balear se presentan en la Tabla 1.
Tabla 1. Éxito de eclosión y éxito reproductor de la pardela balear en seis colonias de 1986 a 2004. Los tamaños de muestra (número de nidos muestreados) se señalan entre paréntesis (Louzao et al., 2006a).
Estructura y dinámica de poblaciones Análisis demográficos recientes han constatado el declive de la Pardela Balear con una disminución anual de la tasa de crecimiento poblacional de un 7.4% (Oro et al., 2004). Al mismo tiempo, han permitido determinar los parámetros demográficos que afectan negativamente a la tasa de crecimiento poblacional: la mortalidad adulta es la principal causa del declive poblacional, mientras que los valores actuales de éxito reproductor (otro parámetro demográfico de potencial relevancia para la conservación de la pardela balear) no son cuestiones preocupantes para la conservación de la especie, al menos en la actualidad y en las colonias de estudio (Oro et al., 2004; Louzao et al., 2006a). La supervivencia adulta estimada fue anormalmente baja (0.780, SE = 0.020, 95% intervalo de confianza: 0.739-0.816) para una especie longeva como la pardela balear, que debería de caracterizarse por una supervivencia adulta mayor (mayor del 0.900) (Oro et al., 2004). También se realizó un análisis de viabilidad de la población mostrando que las probabilidades de extinción para la especie eran extremadamente altas, estimándose que la especie podría extinguirse aproximadamente en 40 años (SE = 0.2) (Oro et al., 2004). Partiendo de una población entre 1.750-2.125 pp, en 54 años (estima de tres generaciones) quedarían 29-36 pp. La modelización para estimar las probabilidades de extinción se basó en las estimas de parámetros demográficos disponibles para la especie (supervivencia por edades, edad a la primera reproducción, probabilidad de reproducirse-años sabáticos y fecundidad); de aquellas de las que no se dispone de estimas se fijaron los valores más altos registrados nunca para especies similares. La fiabilidad de las previsiones se puede suponer alta dado que el parámetro más sensible del modelo, la supervivencia adulta, se estimó de manera robusta por captura-recaptura (Oro et al., 2004). El origen del declive, asociado a la supervivencia adulta, no estaba ligado a problemas de conservación en las colonias de cría, ya que las colonias estudiadas no presentaban depredadores introducidos (gracias a la erradicación de los mismos). Por tanto, la mortalidad de individuos adultos debía provenir principalmente de las zonas de alimentación, es decir, del mar. Por mucho que se mejore la fecundidad, si no se frena la mortalidad adulta en el mar es imposible detener el declive poblacional que predicen los modelos (ver Fig. 1).
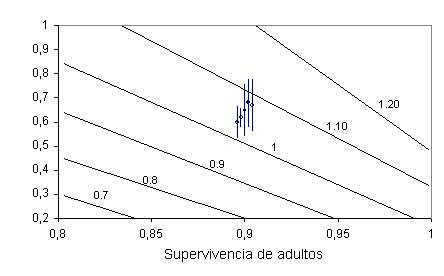
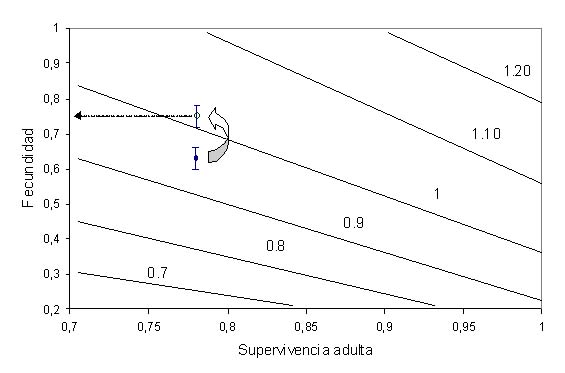
Figura 1. Fitness landscape de supervivencia adulta frente al éxito reproductor, trazados sobre curvas de diferente valor de tasa de crecimiento poblacional l (l = 1 representa la curva de estabilidad de la población). En la figura superior se muestran los valores promedio de éxito reproductor (junto al 95% de intervalo de confianza) de 5 colonias de estudio con datos suficientes, asumiendo una supervivencia adulta típica de 0.9. En la figura inferior se representa el valor actual de estima de supervivencia para la especie (0.78 de Oro et al. 2004, mostrado por la barra de error con marcador sólido) frente al éxito reproductor medio de 5 colonias de estudio (con su 95% de intervalo de confianza); la flecha señala un valor hipotético de éxito reproductor (mostrado por la barra de error con marcador vacío) necesario para que la población se estabilice o crezca (l > 1). Las barras de error son intervalos de confianza del 95% (Louzao et al., 2006a)
Un estudio más reciente, realizado durante el periodo 1985-2014 en la cueva de Sa Cella (Mallorca), mostró que el reclutamiento tuvo lugar sobre todo a los seis años de edad, aunque comenzó a los tres años de edad. El éxito reproductivo medio fue de 0.665, variando entre 0.400 y 0.920 volanderos por pareja. El éxito reproductivo se correlacionó positivamente con la disponibilidad de descartes de pesca. La supervivencia de los adultos se estimó en 0.809 y se debió sobre todo a la mortalidad en artes de pesca (0.455). Este estudio predice un tiempo para la extinción en 61 años (intervalo de confianza 95%: 55-69) (Genovart et al., 2016)1. Al igual que el resto de las Procellariiformes presenta tiempos medios de generación altos (sobre 20 años), supervivencia adulta alta (a pesar de su declive), reclutamiento progresivo y lento, y fecundidad baja (Arcos y Oro, 2004; Oro et al., 2004). Las tasas de dispersión, la reproductora principalmente, parecen bajas (Aguilar, 2000), aunque se tienen ya evidencias genéticas de que la dispersión natal es más alta (Genovart et al., 2007). Existe pues una débil estructuración poblacional fruto de una dispersión mucho mayor de lo que se suponía hasta hace bien poco tiempo (Genovart et al., 2007). Los estudios genéticos también han revelado que hay heterogeneidad en la calidad del hábitat de cría y que algunas colonias sólo parecen mantenerse por flujo de inmigrantes (Genovart et el., 2007). Esta misma conectividad es la que puede haber causado la introgresión de pardela mediterránea P. yelkouan en las colonias de Menorca (Genovart et al., 2005). La estructura poblacional y la heterogeneidad en la calidad de la colonia genera diferencias demográficas importantes en colonias cercanas sometidas a condiciones ambientales parecidas (Tavecchia et al., 2008).
Referencias Aguilar, J. S. (2000). La población de Pardela Balear (Puffinus mauretanicus) en el Parque Nacional del archipiélago de Cabrera. Pp. 33-44. En: Pons, G. X. (Ed.). Las aves del Parque Nacional marítimo-terrestre del archipiélago de Cabrera (Islas Baleares, España). GOB-Ministerio de Medio Ambiente, Madrid. Arcos, J. M., Oro, D. (2004). Pardela Balear, Puffinus mauretanicus. Pp. 46-50. En: Madroño, A., González, C., Atienza, J. C. (Eds.). Libro Rojo de las Aves de España. Dirección General para la Biodiversidad-SEO/BirdLife, Madrid. Genovart, M., Arcos, J. M., Alvarez, D., McMinn, M., Meier, R., Wynn, R. B., Guilford, T., Oro, D. (2016). Demography of the critically endangered Balearic shearwater: the impact of fisheries and time to extinction. Journal of Applied Ecology, 53 (4): 1158-1168. Genovart, M., Juste, J., Oro, D. (2005). Two sibling species sympatrically breeding: a new conservation concern for the critically endangered Balearic shearwater. Conservation Genetics, 6: 601-606. Genovart, M., Oro, D., Juste, J., Bertorelle, G. (2007). What genetics tell us about the conservation of the critically endangered Balearic Shearwater? Biological Conservation, 137: 283-293. Genovart, M., Louzao, M., Igual, J. M., Oro, D. (2008). Digit length may reveal unusual breeding behaviour in a seabird. Biology Letters doi:10.1098/rsbl.2008.0272 Louzao, M., Igual, J. M., McMinn, M., Aguilar, J. S., Triay, R., Oro, D. (2006a). Breeding performance of the critically endangered Balearic Shearwater: improving the conservation diagnosis. Marine Ecology Progress Series, 318: 247-254. Louzao, M., Arcos, J. M., Hyrenbach, K. D., Abelló, P., Gil De Sola, L., Oro, D. (2006b). Oceanographic Habitat of the Critically Endangered Balearic Shearwater: Identifying Suitable Marine Protected Areas. Ecological Applications, 16: 1683-1695. Louzao, M., Igual, J. M., Genovart, M., Forero, M. G., Hobson, K. A., Oro, D. (2008). Inter-population variation in egg size of an endangered Procellariiform: interplay between body size and feeding ecology. Acta Oecologica, 34: 186-193. Oro, D., Aguilar, J. S., Igual, J. M., Louzao, M. (2004). Modelling demography and extinction risk in the endangered Balearic shearwater. Biological Conservation, 116: 93-102. Ruiz, A., Martí, R. (Eds.) (2004). La pardela balear. SEO/BirdLife-Conselleria de Medi Ambient del Govern de les Illes Balears. Madrid. Tavecchia, G., Minguez, E., de León, A., Louzao, M., Oro, D. (2008). Living close, doing differently: small-scale asynchrony in demographic parameters in two species of seabirds. Ecology, 89: 77-85.
Daniel Oro1, Maite Louzao1, 2 y Meritxell Genovart1 Fecha de publicación: 22-06-2009 Otras contribuciones: 1. Alfredo Salvador. 22-09-2016 Oro, D., Louzao, M., Genovart, M. (2016). Pardela Balear – Puffinus mauretanicus. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Morales, M. B. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||