|
|
||||||||||||||||||||
|
||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Purple Swamphen, reproduction, demography.
Biología de la reproducción El modo de reproducción habitual es la cría en parejas. También se han detectado intentos de cría en grupo, algunos de ellos culminados con éxito (Sánchez-Lafuente, 1993; J. Sargatal, com. pers.). Nido El nido del Calamón es una estructura simple (Figura 1), construida con la propia vegetación del medio, y generalmente compuesta de dos partes: una principal, el nido propiamente dicho, en el que las hembras deposita los huevos y que no es sino una depresión en una plataforma de vegetación aplastada de mayor o menor extensión; y otra accesoria, que puede estar presente o no, la rampa de acceso a la plataforma. Esta rampa suele aparecer en nidos construidos en determinados tipos de vegetación (enea Thypha sp. y carrizo Phragmites sp.), probablemente debido a que la elevada densidad de pies por metro cuadrado que caracteriza a estas especies palustres y que dificulta el acceso de los reproductores a la plataforma. Dicha rampa, además se va ensanchando con el continuo trasiego de las aves entrado y saliendo de la plataforma. La rampa, sin embargo, puede faltar en construcciones realizadas sobre vegetación de menor porte o menor densidad. El nido (n= 21) mide en promedio 33 cm de diámetro externo (rango= 27-50 cm), 20,9 cm de diámetro interno (rango= 13-32 cm), 4,2 cm de profundidad de la copa (rango= 1-8 cm) (Grussu, 1999)1. El diámetro medio externo del nido en Argelia fue 34,1 cm, el interno 22,1 cm y la altura 27,8 cm (n= 44) (Samraoui et al., 2015)2.
Figura 1. Nido de calamón. (C) A. M. Sánchez-Lafuente
El Calamón no construye un único nido en su territorio, sino un número variable de ellos. Finalmente, de entre todas las construcciones elige una par hacer la puesta. De hecho, aproximadamente un 70% de los nidos construidos son finalmente rechazados como lugares definitivos para la puesta (Sánchez-Lafuente, 1992), aunque puede ser usados como lugares alternativos para segundas puestas o puestas de reemplazamiento. La construcción finalmente elegida no es necesariamente la más protegida por la vegetación que la rodea, sino la más próxima a un área libre de vegetación, si bien de entras las más próximas a áreas abiertas suelen escoger la de mayor protección (Sánchez-Lafuente et al., 1998). La distancia mínima entre nidos ocupados es de 4 m en Cerdeña (Grussu, 1999)1. Fenología de puesta El periodo reproductor del Calamón común puede variar en función de las condiciones del medio, tanto ecológicas como climatológicas. Así, mientras que en algunas áreas permanentemente inundadas del sur de España puede criar durante todo el año (J. A. Amat, com. pers.), en zonas más septentrionales del interior peninsular (Jaén) sólo se reproduce entre febrero y septiembre, aún cuando las condiciones de inundación sean adecuadas (Sánchez-Lafuente, 2004). En Cerdeña se han registrado puestas todos los meses del año, con máximos en diciembre-enero y marzo-mayo (Grussu, 19991). La incubación dura entre 24 y 34 días (promedio = 29 días, N = 166 huevos, Sánchez-Lafuente, 2004). Tamaño de puesta El tamaño de puesta oscila entre 2 y 8 huevos en la mayoría de los estudios en las distintas áreas de distribución (Hidalgo, 1973; Cramp, 1980; Bauer et al., 1982; Sánchez-Lafuente, 1992; Ferrer et al. 1986), con un valor medio estimado de 4 huevos por nido (Figura 1). Las hembras suelen poner un huevo cada día, si bien en puestas de 6-7 huevos se ha observado que el último, y a veces el penúltimo, huevo se depositaba con una diferencias de hasta 48 horas (Sánchez-Lafuente, obs. Pers.). El tamaño medio de puesta registrado en los humedales de Garaet Hadj Tahar y Garaet Messaoussa (Argelia) fue de 2,75 huevos (Mouslim et al., 2014)2. El tamaño medio de puesta varió entre años en Argelia entre 3,4 y 5,8 huevos (Samraoui et al., 2015)2. Sobre tamaño y peso de los huevos, ver Tabla 1 y Figura 2. El tamaño medio de los huevos en Cerdeña es de 57,6 x 37,2 mm (n= 58) (Grussu, 1999)1. El tamaño medio de los huevos medidos en Argelia fue de 55,2 x 36,6 mm (n= 36) (Samraoui et al., 2015)2.
Tabla 1. Medidas y volumen (± error estándar) de huevos de Calamón común en el Alto Guadalquivir, para puestas de 3, 4 y 5 huevos. Según Sánchez-Lafuente (1992).
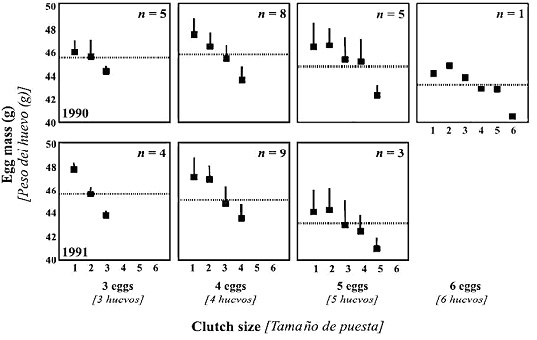
La disponibilidad de alimento alrededor del nido es el factor más importante para determinar la variación en el tamaño de puesta y el peso del huevo. La variación en estas condiciones puede hacer que el tamaño de puesta y el peso del huevo están negativamente relacionados, de forma que el peso medio del huevo disminuye conforme aumenta el tamaño de puesta (Figura 2).
Figura 2. Peso medio (+ error estándar) de huevos de Calamón común en el Alto Guadalquivir, para puestas de 3 - 6 huevos. Según Sánchez-Lafuente (2004).
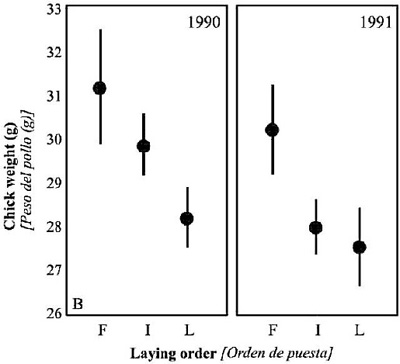
Además, dentro de un mismo nido, los huevos más tardíos suelen ser de menor tamaño, y pueden tener, por tanto, menos probabilidades de eclosionar (Figura 3).
Figura 3. Masa corporal (± error estándar) de los pollos de Calamón común en el Alto Guadalquivir, según su orden de puesta (F = primero, I = intermedio, L = último). Según Sánchez-Lafuente (2004).
En Argelia se han registrado diferencias en el peso y tamaño de los huevos entre sitios y entre años (Mouslim et al., 2014)2. Los pollos pueden considerarse semi-precoces, pues son capaces de abandonar el nido tras eclosionar durante breves períodos no superiores a 24 horas y son capaces de alimentarse por si mismos a los 5-6 días de eclosionar (Sánchez-Lafuente, 2004).
Estructura y dinámica de poblaciones El éxito medio de nacimientos registrado en los humedales de Garaet Hadj Tahar y Garaet Messaoussa (Argelia) fue del 51% y se correlacionó positivamente con la profundidad del nido en Garaet Hadj Tahar (Mouslim et al., 2014)2. La tasa de nacimientos registrada en Boussedra y Tonga (Argelia) varió entre el 82% y el 100% (Samraoui et al., 2015)2. Hay elevada mortalidad de los pollos en Cerdeña (Grussu, 1999). El 40% de los pollos mueren durante los primeros seis días de edad, volando en promedio 1,23 pollos/nido en Jaén (Sánchez-Lafuente, 1992)1. El éxito de nidificación registrado en nidos en los que había al menos un pollo hasta su vuelo, varió en Argelia entre el 56,3% en Tonga y el 76,2% en Boussedra (Samraoui et al., 2015)2.
Referencias Glutz von Blotzheim, U. N., Bauer, K. M., Bezzel, E. (1973). Handbuch der Vögel Mitteleuropas. Band 5. Galliformes und Gruiformes. Aula Verlag, Wiesbaden. Cramp, S. (Ed.) (1980). Handbook of the Birds of Europe, the Middle East and North Africa. The Birds of the Western Palaearctic. Volume II. Hawks to Bustards. Oxford University Press, Oxford. Ferrer, X., Vilalta, A., Muntaner, J. (1986). Ocells. Historia natural dels Països Catalans. Vol. 12. Enciclopedia Catalana. Barcelona. 455 pp. Grussu, M. (1999). Status and breeding ecology of the purple swamp-hen in Italy. Bristish Birds, 92: 183-192. Hidalgo, J. (1973). El calamón común. Vida silvestre, 5: 44-48. Mouslim, B., Eddine, M. S., Rassim, K., Zihad, B., Moussa, H. (2014). Aspects of the breeding ecology of the Purple Swamphen Porphyrio porphyrio in the wetland complex of Guerbes-Sanhadja, north-east Algeria. Ostrich, 85 (2): 185-191. Samraoui, F., Nedjah, R., Alfarhan, A. H., Samraoui, B. (2015). An overview of the Rallidae of Algeria with particular reference to the breedind ecology of the Purple Swamp-Hen Porphyrio porphyrio. Wetlands Ecology and Management, 23 (3): 505-517. Sánchez-Lafuente, A. M. (1992). Biología reproductora de Porphyrio porphyrio en el Alto Guadalquivir. Tesis Doctoral. Universidad de Granada. Sánchez-Lafuente, A. M. (1993). Breeding systems related to incubation investment in the Purple swamphen Porphyrio porphyrio spp porphyrio. Ardea, 81: 121-124 Sánchez-Lafuente, A. M. (2004). Trade-off between clutch size and egg mass, and their effects on hatchability and chick mass in semi-precocial purple swamphen. Ardeola, 51: 319-330. Sánchez-Lafuente, A. M., Alcántara, J., Romero, M. (1998). Nest-site selection and nest predation in the Purple swamphen (Porphyrio porphyrio). Journal of Field Ornithology, 69: 563-576.
Alfonso M. Sánchez-Lafuente Fecha de publicación: 23-03-2010 Otras contribuciones. 1. Alfredo Salvador. 30-11-2016; 2. Alfredo Salvador. 24-03-2017 Sánchez-Lafuente, A. M. (2017). Calamón – Porphyrio porphyrio. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Morales, M. B. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|||||||||||||||||||
|
|
||||||||||||||||||||
|
|
||||||||||||||||||||