|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Eurasian Jackdaw, breeding success, clutch size, daily activity pattern, dispersion, egg colour, egg losses, egg shape, egg size, incubation, laying time, nest building, nest building materials, nest site, nestling mortality, nestling period, productivity, reproduction, testicles development.
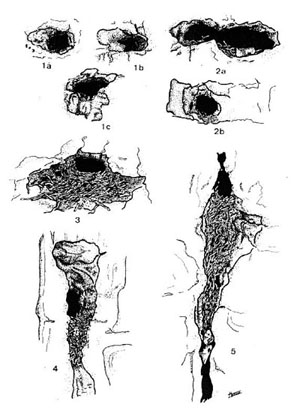
Biología de la reproducción La Grajilla es una especie monógama, tanto a nivel social como a nivel genético (Henderson y Hart, 1993), en la que se mantienen las parejas durante varios años, frecuentemente durante toda la vida (Lorenz, 1931; 1982; Röell, 1978). La biología de la reproducción de la Grajilla es bien conocida. Aunque se han estudiado pocas poblaciones, algunas de ellas se han estudiado en profundidad. En España se han estudiado tres poblaciones, una en la comarca de Guadix (provincia de Granada) (Soler, 1984), otra en las proximidades de la ciudad de León (Rubio Argüello y Ena Álvarez, 1986) y otra en la provincia de Madrid (Domínguez, 1999). Curiosamente estos dos últimos estudios se realizaron durante los mismos años: 1984 y 1985, habiéndose llevado a cabo el primero entre 1979 y 1983. La Grajilla, tiene una clara tendencia a nidificar en colonias, aunque el número de parejas es muy variable dependiendo de la disponibilidad de cavidades apropiadas para la instalación del nido. Este es el caso de la población de la provincia de Granada, donde construyen el nido en cavidades situadas en los taludes naturales. En esta población se puede hablar de nidificación semicolonial ya que abundan los nidos solitarios o en grupos pequeños. La media del número de nidos en las 8 colonias que incluían 10 o más nidos fue de 19,9 (SD = 11,3), siendo la colonia más numerosa una situada cerca de Brácana, en un talud fluvial con 41 nidos (Soler 1984). Parece que esta reproducción semicolonial es lo más frecuente en España. Según De Juana (1980), en la Rioja, lo más normal son colonias de 6-12 parejas o menos. Arroyo (1977) cita una enorme concentración de 2500 parejas nidificantes en un cantil de 6600 m. de longitud situado en la provincia de Madrid. Domínguez (1999) no estudia el número de nidos por colonia, pero encuentra, en la provincia de Madrid, 129 colonias en las que crían 2736 parejas, por tanto, nidifican una media de 21,2 parejas de Grajilla por colonia. El nido: orientación y soporteEl estudio más completo sobre el nido de la Grajilla es el realizado en la provincia de Granada, principalmente en la comarca de Guadix, basado en un total de 1710 nidos que ocuparon un total de 1001 cavidades diferentes (Soler, 1984). (Ver apartado selección de hábitat para información sobre el emplazamiento del nido en diferentes poblaciones). La orientación del nido se estudió en 200 cavidades diferentes correspondientes a nidos localizados en la Hoya de Guadix. La orientación seleccionada con más frecuencia fue noreste (35,5% de los nidos), seguida de norte (19,0%), sur (15,0%) y este (12,5%) (Soler y Soler, 1987). La altura de los nidos se midió en 203 cavidades, observándose un predominio de las situadas entre 2 y 10 metros de altura (media = 7,7 m; SE = 0,3) (Soler y Soler, 1987). La cavidad que soporta el nido (el soporte), es una bolsa de muy diversas formas. Soler y Soler (1987), en un estudio sobre 860 nidos consideran 5 tipos distintos de soporte: (1) cavidades estrechas sin ramas observables desde el exterior (44,9%), (2) cavidades con ramas próximas a la entrada (30,0%), (3) cavidades de entrada horizontal estrechada mediante ramas (8,7%), (4) cavidades de entrada vertical estrechada mediante ramas (5,8%), y (5) grietas (10,6%) (ver Figura 1). Estos resultados ponen de manifiesto que la Grajilla selecciona cavidades de entrada estrecha (tipos 1 y 2), y que cuando la entrada es ancha, la estrechan utilizando ramas que, en los tipos 3, 4 y 5 quedan muy bien encajadas, constituyendo la muralla (ver más abajo apartado “estructura del nido” que dificulta el acceso a los depredadores aunque no siempre resulta eficaz (Soler, 1989a). En 7 cavidades en las que se consiguió medir ancho y alto de la entrada antes y después de la construcción del nido, se obtuvo que tanto el ancho como el alto eran significativamente más grandes antes (media = 35,1 cm; SE = 10,05 para el ancho y media = 23,6 cm; SE = 2,27 para el alto) que después (media = 16,8 cm; SE = 1,08 para el ancho y media = 11,0 cm; SE = 1,54 para el alto).
Figura 1. Tipos de soporte utilizados por la Grajilla en la Hoya de Guadix. Según Soler y Soler (1987a).
Se midieron las dimensiones de los huecos (ancho, alto y profundo) en un total de 109, 105 y 82 cavidades respectivamente, obteniéndose los siguientes resultados: ancho = 19,6 cm (SE = 1,24), alto = 19,2 (SE = 0,8) y profundo = 47,4 (SE = 1,73). Las cavidades utilizadas por las grajillas en la comarca de Guadix, en su mayoría son de origen natural (erosión), pero bastantes son antiguas madrigueras de conejo que, al producirse un corte en el terreno quedan situadas en un talud. Algunas, son antiguos nidos de Pito Real, que en esta zona, con relativa frecuencia, los excava en los taludes de arcilla en lugar de en los troncos de los árboles (Soler et al., 1982). Se han citado otros tipos de soporte utilizados por las grajillas, por ejemplo, cavidades excavadas por el Martín Pescador (Mead y Peplar, 1975); cajas de anidar del tamaño adecuado (Folk, 1968); nidos antiguos de Cigüeña (Ejgelis, 1958; Domínguez, 1999), Urraca, Graja, Corneja y Garza Real (Holyoak, 1967). También se han descrito nidos dispuestos libremente sobre las ramas de los árboles presentando una cúpula, similar a la que tienen los de Urraca, en Gran Bretaña (Owen, 1931; Holyoak, 1967), Polonia (Kulczycki, 1973), Holanda (Röell, 1978) y, también, España, concretamente en el Parque Nacional de Doñana (Álvarez y Arias de Reyna, 1974). Se ha observado en Cataluña una pareja de grajillas aportando material a un antiguo nido de urraca (Geroudet, 1955)1. En algunas ocasiones se ha observado que las grajillas pueden agrandar o incluso construir completamente la cavidad en material blando, en una cantera de yeso (Quantz, 1930) o en troncos con madera podrida (Owen, 1930). Soler y Soler (1987) no observaron en ningún caso la manipulación de la cavidad, lo que parece ser lo más frecuente (Folk, 1968; Kulczycki, 1973). Dimensiones y estructura del nidoSoler (1987) estudió las dimensiones de 21 nidos que se consiguieron sacar de la cavidad completos y sin deformar. Obtuvo los siguientes valores: diámetro de la taza = 12,2x13,3 cm (SD = 0,98-1,09); profundidad de la taza = 5,8 cm (SD = 0,79); Diámetro del nido = 26,1x34,6 cm (SD = 5,86-19,24); espesor del nido = 5,94 cm (SD = 6,66) y, altitud de la cavidad = 11,3 m (SD = 7,57). Las dimensiones correspondientes a la taza eran bastante similares en todos los nidos lo que es lógico puesto que están en relación con el tamaño del ave, pero, las dimensiones del nido eran enormemente variables dependiendo del tipo de soporte (información más detallada de las dimensiones de los nidos dependiendo del tipo de soporte se pueden consultar en Soler (1987)). La altura y el espesor del nido son los parámetros más variables, sobre todo cuando el soporte son grietas, debido a que la base de ramas la hacen tan grande como sea necesario para rellenar la grieta. El espesor va a depender del espacio disponible en la cavidad, en las cavidades muy estrechas no hay ramas, mientras que en las muy amplias las hay en grandes cantidades. Como consecuencia de esto el peso de los nidos también es enormemente variable, entre 13 y 3760 g siendo el valor medio de 516,1 g (SD = 68,6). La mayoría de los nidos, el 52,4%, presentaban un peso inferior a 300 g. La estructura de los nidos también es enormemente variable según el tipo y las dimensiones de la cavidad. Soler (1987) distinguió siete tipos diferentes dependiendo de las capas existentes. La estructura más simple estaría formada por sólo una capa, el forro externo, sobre la que se deposita la puesta, y se puede ir complicando hasta nidos con cuatro capas: forro externo, forro interno, pared y muralla. Forro externo y forro interno son capas estrechas formadas por materiales blandos (los del forro interno algo menos), mientras que la pared y la muralla están formadas principalmente por ramas. La pared es la tercera capa que rodea la taza y la más variable de todas, puede no existir y, cuando existe, su tamaño oscila ampliamente dependiendo de la magnitud de la cavidad. La muralla no rodea uniformemente al nido, sino que recubre a las otras capas en la parte orientada hacia la entrada y se proyecta hacia ésta estrechándola. Información más detallada sobre los distintos tipos de estructura, las capas del nido, las formas de los nidos y cómo se disponen en la cavidad se puede consultar en Soler (1987). Materiales utilizados en la construcción del nidoSoler y Soler (1987b) realizaron un detallado estudio sobre los materiales utilizados por las grajillas en la construcción del nido en la comarca de Guadix (provincia de Granada) basado en el análisis de 108 nidos que fueron recogidos al final del periodo reproductor, entre los años 1980 y 1984. Los elementos utilizados más frecuentemente fueron lana, existente en el 96,3% de los nidos, esparto (91,7%), papel (82,4%), corteza de árbol (80,5%), ramitas (63,9%), pelo (59,2%) y colillas de cigarro 58,3%). Los materiales utilizados varían enormemente según el tipo de soporte en el que está instalado el nido, principalmente el número y el tamaño de las ramas. Se comprobó que cuanto mayor es la cavidad y el tamaño de la entrada, mayor es el número, el tamaño de las ramas y la proporción de ramas bifurcadas, lo que consigue una mayor solidez de la estructura. Harrison (1960) observó 3 grajillas que arrancaban mechones de pelo de un asno que luego transportaban a sus nidos en el castillo de Fuengirola (Málaga)1. Llama la atención la elevada frecuencia de aparición de las colillas de cigarro, presentes en el 58,3% de los nidos, sobre todo teniendo en cuenta que las colonias estudiadas se encuentran bastante retiradas de vertederos de basura, aunque algunas de ellas si están próximas a alguna carretera. En 11 de los nidos había más de 10, llegando a 43 en el caso máximo. Parece claro que existe una predilección por las colillas en las grajillas de la comarca de Guadix. Este elemento no ha sido citado en la bibliografía como componente de los nidos de Grajilla, pero sí otros igualmente artificiales y de escaso valor estructural como trozos de goma o de metal. El caso más llamativo es el de las pinzas de la ropa. Kirchner (1933) encontró en una población una clara predilección de las grajillas por las pinzas de la ropa encontrando hasta 58 en un nido. Observó que las aves llegaban a cogerlas directamente de las cuerdas de tender. Con frecuencia (en el 75% de los nidos), sólo uno de los elementos constituía más del 50% del volumen de materiales. Cuando la cavidad es amplia ese elemento mayoritario son las ramas y, cuando es pequeña, es un material blando, principalmente lana, pero también, en algunos nidos, fue el pelo. Es frecuente la existencia de ramas en la base de los nidos, seguramente han sido desechadas o se les han caído cuando intentaban meterlas en la cavidad. Otra posible explicación basada en descubrimientos más recientes, es que el transporte de ramas y otros elementos pesados forme parte de una exhibición de fuerza, por parte del macho a la hembra (Soler et al. 1998). Sería una señal que permitiría a la hembra ajustar su esfuerzo reproductor durante esa temporada. Este argumento basado en la teoría de la selección sexual está apoyado por el hecho de que, con frecuencia, son abundantes las ramas debajo de los nidos situados en cavidad pequeña en los que no es posible incluir ninguna rama (Soler y Soler 1987b). Los materiales para la construcción del nido, normalmente, se asume que se utiliza el material adecuado disponible en las proximidades de los nidos, ya que el transporte es una actividad muy costosa y hay un conflicto importante de economía de esfuerzo (Collias y Collias, 1884). Hay muy poca información sobre este tema pues las observaciones publicadas prácticamente siempre se realizan en las proximidades de los nidos. En la Grajilla, para 27 observaciones de recogida y transporte de una rama al nido, la distancia media fue de 16,4 m (SE = 2,4) (Soler y Soler, 1992a). Los materiales blandos, imprescindibles para la taza, son más difíciles de encontrar y menos pesados, por lo que se asume que se buscarán y se transportarán desde mayores distancias. Soler y Soler (1992a) realizaron un experimento para determinar la distancia a la que las grajillas recogen los materiales. Se tiñeron 15 Kg. de borra de cinco colores diferentes y se distribuyeron en cinco montones, uno de cada color, siguiendo una línea recta, a 250 metros cada uno del siguiente. Una vez finalizado el periodo reproductor se recogieron 18 nidos de distintas colonias y se calcularon las distancias de cada uno de los nidos a cada uno de los montones. Los resultados mostraron que el transporte se realiza preferentemente desde menos de 250 m. A distancias entre 250 y 500 m también se realizaron significativamente más transportes que desde distancias mayores, siendo estos últimos ya más escasos. Se detectó, como distancia máxima, transporte de borra a un nido a más de 2000 metros. Coombs (1978) destaca que la Chova Piquirroja puede colectar lana incluso a 2 km del nido lo que coincide con el dato mencionado. Comportamiento de construcción del nidoLa construcción del nido es un proceso complejo en el que normalmente intervienen los dos miembros de la pareja (Collias y Collias, 1984). En el caso de la Grajilla, macho y hembra colaboran tanto en el transporte (Lorenz, 1931; Soler y Soler 1991b) como en la ordenación de los materiales (Zimmermann, 1951; Soler y Soler, 1991b). Con respecto al papel de ambos sexos en la construcción del nido, Soler y Soler (1991b) destacan que: (1) Al principio sólo transporta materiales la hembra. (2) Posteriormente transportan los dos pero principalmente la hembra (69%, n = 36). (3) El macho suele aportar los elementos de mayor tamaño (hecho también destacado por Lorenz (1931)). (4) La ordenación de materiales, cuando la cavidad es estrecha, sólo la realiza la hembra (observado en 8 nidos), pero si la cavidad es amplia la realizan los dos (observado en 16 nidos), permaneciendo la hembra significativamente más tiempo que el macho en el interior del nido (hembra = 272 s (n = 14), macho = 192 s (n = 9)).
Figura 2. Hembra de Grajilla con lana para la taza del nido.
Las parejas más adelantadas comienzan el transporte de materiales durante la última semana de marzo y, sólo ocho días más tarde, ya están en construcción el 50% de los nidos (Soler y Soler, 1991b). La construcción del nido dura, por regla general, entre 18 y 26 días (media = 21,7 días, n = 39). La construcción más rápida se realizó en 8 días. Son varios los factores que pueden influir en la duración del periodo constructor, los más importantes son la temperatura y la fecha de inicio de la construcción, habiéndose observado que una disminución de la temperatura ralentiza la actividad constructora y, un inicio tardío, acelera el proceso (Soler y Soler, 1991b). La actividad diaria de transporte de materiales presenta dos picos muy claros, uno por la mañana (entre las 6:30 y las 10h, hora solar) que comienza después de dedicar la primera hora a comer, y otro por la tarde, más marcado, entre las 14 y las 16h) (Soler y Soler, 1991b). El robo de materiales blandos es bastante frecuente en las colonias de grajillas. El material robado no siempre es incluido en el nido del ladrón (sólo en 11 de 31 observaciones) (Soler y Soler, 1991b). Hay parejas que se especializan en robar materiales y, en este caso, sí los incluyen siempre en su nido. Soler y Soler (1991b) citan un caso de una pareja especialista: de 17 aportes de material blando observados en diferentes días, 13 fueron robados, tres recogidos que se les cayeron a sus vecinos y sólo uno de los aportes fue traído desde una distancia indeterminada. Esto supone 13 robos de 17 aportes de material frente a sólo 6 robos de 46 aportes observados en las cuatro parejas vecinas. También se ha citado a la Grajilla robando material de nido a otras especies como los buitres (Elósegui, 1985). Desarrollo de las gónadasEn aves, está ampliamente aceptada la idea de que el aumento paulatino del fotoperiodo es lo que provoca el desarrollo de las gónadas que, a su vez, es el responsable de que se produzcan las hormonas que provocan los diversos comportamientos reproductores. Se estudió el tamaño de los testículos en 84 machos de Grajilla capturados a lo largo de todo el año, y se puso de manifiesto que mantenían un tamaño más o menos constante (entre 3,2 y 4,7 mm) excepto durante los meses de marzo, en el que se observa un aumento del tamaño, y abril y mayo, en los que alcanzan su máximo comenzando a disminuir ya en la segunda quincena de este último mes (Soler, 1984). Tabla 3. Dimensiones de los testículos en machos de Grajilla (mm) entre los meses de marzo a mayo divididos por quincenas.
Estos datos apoyan la idea general mencionada anteriormente, ya que las grajillas de esta población comienzan a construir el nido durante la última semana de marzo (ver más arriba), coincidiendo con el primer crecimiento importante de los testículos, y la puesta, a primeros de mayo (ver más abajo) coincidiendo con el máximo tamaño.
Fenología de puestaEn la comarca de Guadix, el inicio de las puestas comienza, en la mayor parte de los nidos (n = 192), entre el 26 y el 28 de abril (29,2% de las puestas). La más temprana tuvo lugar el 20 de abril y las más tardías a partir del 8 de Mayo. La fecha media de inicio de las puestas fue 28,1 de abril (SD = 4,6) (Soler y Soler 1987c). Información más detallada especificando para los cinco años (1979-1983) y las cuatro zonas de estudio se puede consultar en Soler y Soler (1987c). En León, las puestas se iniciaron en mayo durante los dos años de estudio (media = 16,1 de mayo; n = 64) (Rubio Argüello y Ena Álvarez, 1986). No encuentran diferencias entre años (17,7 y 15,1 de mayo respectivamente para 1984 y 1985). Información más detallada sobre los dos años de estudio, especificando número de nidos que inician la puesta cada semana, se puede encontrar en Rubio Argüello y Ena Álvarez (1986). En la provincia de Madrid, la fecha de puesta osciló entre los últimos días de abril y los primeros de mayo, siendo la mediana para el total de los datos el 5 de mayo (Domínguez 1999). Encuentra diferencias significativas entre años siendo la fecha de puesta más tardía durante 1984 que durante 1985 (mediana = 6 y 2 de mayo respectivamente). La población de León comienza las puestas con un retraso de 18 días respecto a la población granadina, mientras que la población de Madrid es algo más temprana que esta última. Curiosamente, la fecha de puesta encontrada en la zona de Guadix es de las más tardías encontradas en la bibliografía (la correspondiente al año 1981 = 30,6 de abril, es posterior a todas ellas), y la encontrada en León, es, con diferencia la más retrasada de Europa (los datos sobre fecha de puesta en otras poblaciones europeas se pueden consultar en la Tabla 2 de Soler y Soler (1987c)). Esto es bastante extraño teniendo en cuenta que, por regla general, las puestas se inician antes en latitudes más meridionales y, sin embargo, las grajillas comienzan a poner antes en países mucho más septentrionales. (Ver Soler y Soler (1987c) para una detallada discusión de los factores que pueden afectar a la fecha de puesta y propuesta de hipótesis que pueden explicar estas diferencias entre zonas). La fecha de puesta se adelantó significativamente tras el aporte experimental de alimento a dos colonias (Soler y Soler, 1986). Las grajillas normalmente realizan una sola puesta (Cramp y Perrins, 1994). Las puestas de reposición no son raras en córvidos, siendo relativamente frecuentes en la Corneja (Yom-Tov, 1974) y la Urraca (Birkhead, 1991). No obstante, en la Grajilla son muy escasas (Richford, 1978) o inexistentes (Zimmermann, 1951; Folk, 1968). En el estudio realizado en la Hoya de Guadix sólo se detectaron 5 puestas de reposición realizadas por parejas cuyo nido había sido depredado durante la puesta (Soler, 1984).
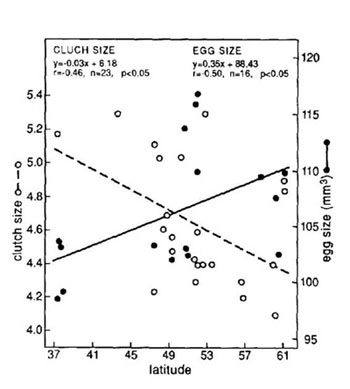
Tamaño de puestaEl tamaño de puesta de la Grajilla oscila entre los 4,1 huevos por nido obtenidos en Leningrado (Ejgelis, 1958) y los 5,3 encontrados en Gales (Holyoak, 1967) y en la Camargue (Gibbons, 1987). Información sobre el tamaño de puesta en diferentes poblaciones de Europa se puede consultar en la Tabla 2 en Soler y Soler (1991c). En España, en las tres poblaciones estudiadas se han encontrado 5,1 huevos por nido (SE = 0,01; n = 173) en la comarca de Guadix (Soler y Soler, 1991c); 4,38 (SD = 1,29; n = 78) en León (Rubio Argüello y Ena Álvarez, 1986) y 5,2 huevos (CV = 0,22; n = 106) en la provincia de Madrid (Domínguez, 1999). Curiosamente, el primer valor es de los más altos mencionados en la bibliografía, y el segundo, de los más bajos. La diferencia entre colonias de una misma población en una estación reproductora no es importante, sin embargo, las diferencias entre años si pueden ser considerables como consecuencia de las variaciones climatológicas que pueden afectar al tamaño de puesta. Rubio Argüello y Ena Álvarez (1986) encontraron una importante diferencia en el tamaño de puesta de su población en León durante dos años consecutivos (3,97 en 1984; SD = 1,22; 4,65 en 1985; SD = 1,28) explicando el bajo tamaño de puesta durante el primer año (es el más bajo de todos los citados en la bibliografía), como la consecuencia de una climatología especialmente adversa. En Madrid fue ligeramente superior en 1985 aunque las diferencias no fueron significativas (Domínguez, 1999). Soler y Soler (1991c), en su estudio sobre el tamaño de puesta de la Grajilla en la Hoya de Guadix, encontraron que: (1) el tamaño de puesta disminuye significativamente conforme avanza la estación reproductora en los cuatro años de estudio, hecho también destacado para la provincia de Madrid (Domínguez, 1999). (2) Existe una variación geográfica en la que la longitud no tiene una influencia significativa pero la latitud sí (Soler y Soler, 1992b); (3) el tamaño de puesta de 7 hembras marcadas fue significativamente alternativo, es decir, si un año era elevado, al año siguiente era pequeño y viceversa; (4) las hembras de primer año pusieron un tamaño de puesta significativamente menor (media = 4,1; SE = 0,3; n = 15) que el correspondiente a la media de la población; y (5) el tamaño de puesta disminuyó en una de las colonias estudiadas conforme disminuyó la densidad de parejas reproductoras. Soler y Soler (1996) encontraron que el aporte experimental de alimento extra produjo un aumento significativo del tamaño de puesta en estas colonias experimentales (6,0) respecto a las colonias control en las que no se aportó alimento extra (5,4). Como se ha mencionado anteriormente, la latitud afecta significativamente al tamaño de puesta en la Grajilla, sin embargo, lo hace en sentido contrario del que se predice teóricamente: la tendencia general en aves es a aumentar conforme aumenta la latitud, pero en Grajilla, el tamaño de puesta disminuye conforme aumenta la latitud. Soler y Soler (1992b) analizan este dato en relación con el tamaño del huevo que precisamente presenta la tendencia contraria: aumenta significativamente con la latitud (ver Figura 3), y proponen una nueva hipótesis que afectaría a especies que crían en cavidades y realizan una sola puesta por temporada: en estas especies, puestas de mayor tamaño son beneficiosas y decrecen con la latitud como consecuencia del compromiso entre tamaño de puesta y tamaño de huevo. En latitudes más norteñas los huevos de mayor tamaño son muy ventajosos por lo que se disminuye el tamaño de puesta para favorecer un mayor tamaño del huevo.
Figura 3. Relación entre tamaño de puesta, tamaño de huevo y latitud en la Grajilla. Según Soler y Soler (1992b).
La hipótesis de la depredación de nidos predice que conforme aumenta el riesgo de depredación debería disminuir el tamaño de puesta (Martín, 1995). Soler y Soler (1993) testaron esta hipótesis analizando los datos de tamaño de puesta, depredación de nidos y éxito reproductor en 128 nidos de Grajilla de la comarca de Guadix. Como indicador del riesgo de depredación se utilizó la más pequeña de las dimensiones de la entrada a la cavidad, puesto que encontraron que la tasa de depredación estaba positivamente correlacionada con esta medida (basta con que una de las dimensiones de la entrada sea lo suficientemente pequeña como para evitar el acceso al depredador). El tamaño de puesta no resultó afectado por el riesgo de depredación, es decir, los nidos de entrada amplia, con elevadas probabilidades de ser depredados, no tenían un menor número de huevos que los de entrada estrecha. En aves, el tamaño de puesta puede ser “determinado” o “indeterminado” (Klomp, 1970). Se habla de tamaño de puesta “determinado” cuando el número de huevos que pone una hembra no varía aunque se le añadan o quiten huevos (“indeterminado”, en el caso contrario). Soler y Soler (1991c) hicieron un experimento en el que, a unos nidos de Grajilla le añadieron huevos, mientras que a otros les quitaron. Estas manipulaciones no tuvieron ningún efecto sobre el tamaño de puesta que fue similar al de los nidos control en los que no se hicieron manipulaciones. Por tanto se concluía que la puesta en esta especie es determinada.
Tamaño del huevoSe han realizado tres estudios sobre las dimensiones de los huevos de Grajilla en España, uno en la comarca de Guadix, otro en las proximidades de la ciudad de León y, un tercero, en la provincia de Madrid. Los valores obtenidos en las tres poblaciones se recogen en la Tabla 4.
Tabla 4. Dimensiones de los huevos de Grajilla en la Hoya de Guadix (Soler, 1988), León (Rubio Argüello y Ena Álvarez, 1987) y provincia de Madrid (Domínguez, 1999). El volumen se calculó, en la zona de Guadix utilizando la fórmula de Coulson (1963: V = πA2LK/6, donde A es la anchura, L es la longitud y K es una constante derivada de una muestra de huevos cuyo volumen se conoce) y en la zona de Madrid por el índice de Hoydt (1979: V = LA2K, donde A es la anchura, L es la longitud y K es una constante que toma el valor de 0,00051).
Además, hay información sobre el tamaño del huevo de la Grajilla en otras tres poblaciones (en Soler y Soler, 1992b): Centro de España (huevos medidos en el Museo Nacional de Ciencias Naturales): 32,07mm x 24,73mm , n = 8; Mures (Jaén): 33,91mm x 24,52mm, n = 31; y Huéneja (Granada): 32,24mm x 24,59mm, n = 18. Los valores obtenidos en la población de León y en la de Madrid son similares a los encontrados en otras poblaciones europeas, pero los de la población de Guadix son más pequeños. Los de las otras tres poblaciones, el tamaño de muestra es demasiado pequeño para discutirlos. La anchura está incluida entre las obtenidas en otras poblaciones europeas, pero la longitud es la más pequeña de todas las citadas en la bibliografía (las referencias bibliográficas sobre las dimensiones de los huevos en distintas poblaciones europeas se pueden consultar en la Tabla 9 de Soler y Soler (1988)). El tamaño de huevo varía con la secuencia de puesta. Al comparar las dimensiones del primer huevo con el resto y las del último huevo con el resto, Soler y Soler (1988) encuentran diferencias significativas en todos los casos (excepto en la comparación de la longitud del último huevo con la del resto) y, estas diferencias, son en el sentido de que los huevos son más pequeños conforme avanza la secuencia de puesta (el primero era más grande que el resto y el último era más pequeño que el resto). También hay una variación del tamaño de huevo según la fecha de puesta, los huevos son más pequeños conforme avanza la estación reproductora, siendo la correlación estadísticamente significativa (Soler y Soler, 1988). Tanto en León como en Guadix se estudió el tamaño del huevo según el tamaño de puesta. Aunque hay una ligera tendencia a aumentar no es significativa (Rubio Argüello y Ena Álvarez, 1987; Soler, 1988a). El aporte experimental de alimento no provocó un aumento significativo del tamaño del huevo (Soler y Soler, 1996).
Coloración del huevoLos huevos de las grajillas son, en general, de color azulado-verdoso pálido, a veces casi blanco, moteados de manchas de color y tamaño variable aunque predomina el marrón oscuro (Soler, 1988b).
Figura 4. Puesta de Grajilla de 4 huevos.
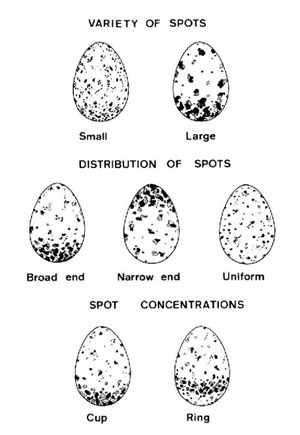
Existen dos estudios en España que analizan cuantitativamente la coloración del huevo de la Grajilla según el tipo y distribución de las motas, uno realizado en la comarca de Guadix (Soler y Soler, 1988), y otro, en las proximidades de la ciudad de León (Rubio Argüello y Ena Álvarez, 1987). Ambos siguen la metodología descrita por Soler (1984) basada en los tipos de huevos representados en la Figura 5.
Figura 5. Tipos de huevos de Grajilla según los patrones que siguen las motas. Según Soler (1984).
Los resultados obtenidos en ambas poblaciones se especifican en la Tabla 5.
Tabla 5. Número y porcentaje de huevos de Grajilla según el tipo, distribución y forma de concentración de las motas en dos poblaciones españolas de Grajilla en la Hoya de Guadix (Soler, 1988b) y en León (Rubio Argüello y Ena Álvarez, 1987). El tipo de moteado “Medio”, y la distribución de moteado “Mitad polo ancho” no fueron considerados en el estudio de la población de Guadix y la distinción entre forma de concentración del moteado no fue tenida en cuenta en la población de León.
Soler (1984) destaca el hecho de que el tipo, la distribución y la forma de concentración del moteado varía para una hembra de un año para otro, e incluso, dentro de una misma puesta, por lo que concluye que las motas no son una característica fijada genéticamente, sino que dependerá del proceso de formación del huevo.
Forma del huevoLa forma del huevo también se ha estudiado en las tres poblaciones y los datos se resumen en la Tabla 6.
Tabla 6. Forma del huevo en las tres poblaciones de Grajilla estudiadas: Guadix (Soler, 1988), León (Rubio Argüello y Ena Álvarez, 1987) y Madrid (Domínguez, 1999). La forma se describe siguiendo (A) la clasificación de Harrison (1977) y (B) el índice de Väisänen (1969), en el que la forma F = 100 x longitud / anchura, siendo el huevo más ovoide cuanto más elevado sea el valor del índice.
La forma del huevo de Grajilla aparentemente se mantiene bastante constante entre poblaciones, pues, además de ser muy similar en las tres poblaciones españolas, también es bastante similar en dos poblaciones de Finlandia (norte: F = 139,4; sur: F = 140,2; Antikainen, 1978).
IncubaciónEn los córvidos, lo normal es que sea la hembra quien se encargue de la incubación (Coombs, 1978). En la Grajilla, también se encarga la hembra que, mientras dura este periodo, es alimentada regularmente por el macho, (Figura 3) (Lockie, 1955; Soler y Soler, 1991b). Cuando la cavidad es amplia la ceba se produce en el interior, pero cuando es estrecha la hembra sale a recibir el alimento.
Figura 6. Macho cebando a la hembra durante el periodo de incubación.
Se trata de una especie de eclosión asincrónica en la que el comienzo de la incubación es muy variable entre nidos. Soler, (1989b), encontró que la Grajilla, en la comarca de Guadix, comienza a incubar, por término medio, antes de la puesta del segundo huevo (media = 1,73, n = 45). Puede empezar con cualquiera de los tres primeros huevos (con el primero en el 48,9% de los casos, con el segundo en el 33,3%, con el tercero en el 13,3%, y con el cuarto, en el 4,4%). Todos los huevos de un mismo nido pueden eclosionar en 2 (n = 2), 3 (n = 18) ó 4 (n = 11) días. El comienzo de la incubación está afectado por el tamaño de puesta. Conforme mayor es el tamaño de puesta más tarde comienza la hembra a incubar los huevos (Zimmerman, 1951; Folk, 1968, Soler, 1989b). El periodo de incubación (desde la puesta del primer huevo hasta la eclosión del primer pollo) fue de 20,8 días (n = 122; SD = 1,85), pero fue bastante variable. Los periodos más frecuentes fueron el de 20 días (22,1% de los casos) y el de 21 (18,9%) (Soler 1989b). El periodo de incubación real (desde la puesta del último huevo hasta su eclosión) pudo determinarse en cinco nidos y fue de 17,2 días (Soler, 1989b).
Actividad durante el periodo de puesta incubaciónSoler y Soler (1991b), realizaron un estudio basado en 153,5 horas de observación abarcando todas las horas del día, durante las que registraron 239 cebas del macho a la hembra. Destacaron los siguientes puntos: (1) las hembras reciben por término medio 1,44 cebas por hora, (2) la ceba dura una media de 83,3 s (SD = 17,2, n = 19). (3) A lo largo del día se detectan dos periodos en los que el aporte de cebas es mínimo: la hora y media posterior al amanecer (debido a que cuando llega el macho la hembra sale del nido y se dedica a comer) y la hora y media que precede al oscurecer. Pérdidas de huevosEl porcentaje de huevos perdidos varía considerablemente entre poblaciones, oscilando entre el 11,5% encontrado por Lockie (1955) y el 28% hallado por Richford (1978). Los valores obtenidos para poblaciones españolas son más elevados (ver Tabla 7).
Tabla 7. Pérdidas de huevos en dos poblaciones españolas de Grajilla, una en la Hoya de Guadix (Soler, 1989a) y otra en las proximidades de León (Rubio Argüello y Ena Álvarez, 1986).
Llama la atención el elevado porcentaje de pérdidas de huevos encontrado en León, principalmente durante 1984, que se debe a un aumento tanto en el número de nidos abandonados como depredados (Rubio Argüello y Ena Álvarez, 1986). Las principales causas de pérdidas de huevos son el fallo en la eclosión, la depredación y el abandono de los nidos (ver Tabla 8).
Tabla 8. Principales factores responsables de las pérdidas de huevos en dos poblaciones españolas de Grajilla, una en la Hoya de Guadix (Soler, 1989a) y otra en las proximidades de León (Rubio Argüello y Ena Álvarez, 1986). Los valores para León recalculados a partir de datos originales asumiendo que los considerados “desaparecidos” por los autores son depredados.
Es de destacar el elevado porcentaje de nidos depredados en León. Los autores destacan que son de fácil acceso, por lo que los principales responsables serían los humanos. En Guadix el principal depredador es el Cuervo, responsable de la depredación de 17 puestas completas que incluían un total de 81 huevos (36,5% de los huevos perdidos). Las hembras jóvenes, reproductoras de primer año, sufrieron unas pérdidas de huevos del 43%, significativamente mayores que las obtenidas para el resto de los nidos (Soler, 1989a).
Periodo de estancia de los pollos en el nidoEl periodo de tiempo transcurrido entre la eclosión y el abandono del nido por parte de los pollos sólo ha sido estudiado en dos poblaciones europeas: Finlandia (33,8 días, n = 75 pollos; Antikainen, 1978) y Hoya de Guadix (Soler, 1988c). En el estudio realizado en Guadix (Soler 1988c) se obtuvo una duración media de 32,4 días (SE = 0,12, n = 109 nidos). El periodo de estancia es bastante variable, habiendo oscilado en el estudio de Guadix entre 28 y 36 días (en el de Finlandia entre 28 y 41 días, Antikainen, 1978). La duración del periodo de estancia de los pollos en el nido está afectada significativamente, tanto por el número de pollos existentes en el nido, como por el número de pollos que llegan a volar, aumentando el periodo de estancia conforme mayor es el número de pollos (Soler, 1988c). El aporte experimental de alimento extra, no sólo no consiguió disminuir el periodo de estancia en el nido, sino que la tendencia fue la contraria (media = 33,2 días en los nidos con alimento extra, 32,6 días en los nidos sin alimento extra) (Soler y Soler, 1996). Este resultado contrario al esperado se debe a que el aporte de alimento provocó un aumento significativo del número de pollos en el nido, lo que produciría el aumento del periodo de estancia.
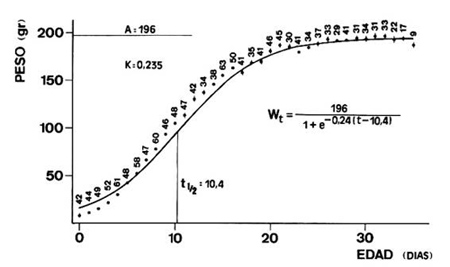
Crecimiento y desarrollo de los pollosEn España sólo se han realizado dos estudios sobre el crecimiento de los pollos de Grajilla, un en la comarca de Guadix (Soler y Soler,1989) y otro en la provincia de Madrid (domínguez, 1999). En la comarca de Guadix los pollos de Grajilla eclosionan con un peso de 7,9 g (SE = 1,8; n = 21). La Figura 7 muestra la curva de crecimiento basada en el aumento de peso. Se observa que los pollos presentan un crecimiento sumamente rápido (durante los primeros 16 días), después continúa más lentamente hasta alcanzar el máximo en el día 31, observándose una pérdida de peso durante los últimos tres días de estancia en el nido.
Figura 7. Crecimiento ponderal de los pollos de Grajilla. Los puntos corresponden a los valores medios, los números indican el tamaño de la muestra y la línea vertical, cuando aparece, representa el error estándar. La curva se ha obtenido ajustando los puntos según el método gráfico de Ricklefs (1967). A es la asíntota, K es la constante proporcional a la razón específica de crecimiento, t50 es la edad a la que se alcanza el 50% de la asíntota y se especifica también la ecuación de crecimiento. El día 0 es el día de la eclosión. De Soler y Soler (1990).
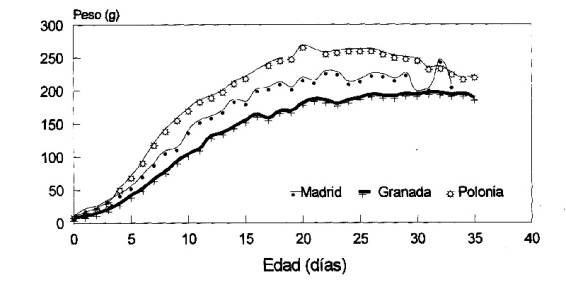
Domínguez (1999), realiza una comparación entre las curvas de crecimiento de los pollos de Grajilla de tres poblaciones europeas (ver Figura 8) y encuentra importantes diferencias entre la población de Guadix y la de Polonia manteniendo la población de Madrid valores intermedios.
Figura 8. Comparación de los valores medios del peso a lo largo del crecimiento en tres poblaciones de Grajilla según datos de Kaminski y Konarzewski (1984) para Polonia, Soler (1984) para Granada y Domínguez (1999) para Madrid. Sacada de Domínguez (1999)
Se estudió el crecimiento tanto de características propias del pollo (comisuras bucales) como del adulto (tarso, ala, cola, etc.) (ver Figura 9). Las primeras crecen muy rápidamente durante los 13 primeros días y después van disminuyendo de tamaño. El tarso alcanza valores similares a los del adulto el día 18 y ya se mantiene.
Figura 9. Registrando las características de dos pollos de Grajilla que están en la fase final del periodo de estancia en el nido.
Los ojos comienzan a abrirse a los 8 días de edad y a los 14 ya están totalmente abiertos en el 90% de los pollos. La piel, que al nacer es de color rojizo, al día siguiente se torna marrón claro y, en los días siguientes se va oscureciendo progresivamente adquiriendo el color negruzco a partir del séptimo día. Los cañones de las remiges comienzan a asomar entre el sexto y el octavo día. Soler y Soler (1990) destacan que se pueden diferenciar dos etapas en el desarrollo de las grajillas que se corresponden con el comportamiento petitorio. Durante la primera se produce un crecimiento muy rápido del peso, tarso y de las características propias de pollo, y coincide con el comportamiento petitorio que durante esa etapa es bastante pasivo, caracterizado porque el pollo simplemente estira el cuello y abre la boca. Este cambia como resultado del desarrollo adecuado de los tarsos, aumento del peso y de la abertura de los ojos. A partir de aquí el comportamiento petitorio es mucho más activo: el pico abierto se dirige hacia la cabeza del adulto y presentan una mayor movilidad. Entonces comienzan a disminuir de tamaño las características propias de pollo y tiene lugar el crecimiento, muy rápido, de las características propias de adulto como ala y cola.
Mortalidad de los pollosLos datos más relevantes sobre la pérdida de pollos en dos poblaciones españolas se presentan en la Tabla 9.
Tabla 9. Pérdidas de pollos en dos poblaciones españolas de Grajilla, una en la Hoya de Guadix (Soler, 1989a) y otra en las proximidades de León (Rubio Argüello y Ena Álvarez, 1986).
Domínguez (1999), encuentra que en Madrid mueren el 61,75% de los pollos que nacen, siendo este porcentaje mayor durante 1984 (77,71%) que durante 1985 (52,51%). La mayor mortalidad detectada durante el primer año fue la consecuencia de las lluvias primaverales que inundaron algunos nidos. En otros estudios realizados en poblaciones europeas se obtienen porcentajes de pérdidas de pollos bastante más bajos (Folk (1968): 33,8%; Antikainen (1978): 36,5%), seguramente debido a que el porcentaje de nidos depredados en estas poblaciones que nidifican en cajas de anidar es considerablemente inferior al encontrado en las poblaciones españolas nidificando en cavidades naturales. En cuanto a los factores responsables de las pérdidas de pollos en las poblaciones españolas, en la de León, los autores sólo diferencian entre “desaparecidos” y “muertos”, siendo los primeros los que desaparecen del nido, y los segundos, los que encuentran muertos en él (Rubio Argüello y Ena Álvarez, 1986). Esta clasificación no permite distinguir entre los pollos que fueron depredados y los pollos muertos por inanición. En la población de Madrid se destaca el efecto negativo de las lluvias primaverales y la caza organizada de grajillas permitidas porque la especie es considerada plaga en la zona. La muerte de uno de los adultos provocaría la muerte por inanición de todos o casi todos los pollos (Domínguez, 1999). En la Hoya de Guadix (Soler, 1989a), la mayor parte de los pollos murieron de inanición (78,0%), siendo las pérdidas debidas a la depredación de un 17.1%. El principal depredador fue, al igual que durante el periodo de puesta-incubación, el Cuervo (8,2%). El hecho de que la falta de alimento sea la responsable de la mayor parte de las pérdidas de pollos se ha destacado en todos los estudios sobre la biología reproductiva de la Grajilla (Zimmermann, 1951; Lockie, 1955; Antikainen, 1978). En esos estudios también se constata que la mayor parte de los pollos mueren durante sus primeros días de vida. En la población de Guadix, el 43,6% de los pollos mueren antes de cumplir los 5 días (Soler, 1989a). Esta elevada mortalidad a una edad temprana que se produce en Grajilla es la consecuencia de que se trata de una especie que sigue una estrategia reproductora conocida como “reductora del número de pollos” (ver apartado “estrategia reproductiva” más abajo). En el estudio de la población de la zona de Guadix (Soler, 1989a) se puso de manifiesto que las condiciones climatológicas tienen un efecto muy importante sobre la mortalidad de los pollos afectando a la disponibilidad de alimento. Se encontró que el número de pollos que moría aumentaba al disminuir las precipitaciones y al aumentar la temperatura. El número de pollos existentes en el nido también afecta a la mortalidad, ya que se encontró que tanto la media como el porcentaje de pollos que mueren aumenta con el tamaño de pollada (Soler, 1989a). La densidad de nidos afecta de forma bastante directa a la tasa de depredación. Johnsson (1994) encontró que conforme aumentaba el tamaño de la colonia disminuía el porcentaje de nidos depredados en una población de grajillas de Suecia donde el principal depredador era la Malta. Soler y Soler (1996) mostraron que tras un tratamiento experimental en el que aportaron alimento extra a dos colonias aumentó considerablemente la densidad reproductora y disminuyó a cero el porcentaje de nidos depredados. En ambos estudios se interpretó esta disminución de la depredación con el aumento del número de parejas como la consecuencia de que las grajillas defendían en grupo los nidos de la colonia.
Productividad
Tabla 10. Éxito reproductor (de eclosión, de volantones y total) y productividad de la Grajilla en tres poblaciones españolas: Hoya de Guadix (Soler, 1990), León (Rubio Argüello y Ena Álvarez, 1986) y Madrid (Domínguez, 1999).
Los valores recogidos en la Tabla 10 son bastante similares a los obtenidos en otras zonas europeas (ver datos y referencias bibliográficas en la Tabla IV, en Soler, 1990). En cuanto a las tres poblaciones españolas, es de destacar que, mientras que el éxito de eclosión es bastante mayor en la población de Guadix, el éxito de volantones es bastante mayor en León, manteniendo la zona de Madrid valores intermedios. El éxito reproductor total y la productividad es bastante similar en las tres zonas (ver Tabla 10). Soler (1990) encontró una relación positiva y significativa entre productividad y densidad de nidos. En un experimento en el que se aportó alimento extra a unas colonias de Grajilla mientras que otras se mantenían como control (sin alimento), el tratamiento provocó un aumento significativo del éxito de volantones y del éxito reproductor total, pero no del éxito de eclosión (Soler y Soler, 1996).
Estrategia reproductivaEn aves se distinguen dos estrategias reproductoras principales: (1) el número de pollos que vuelan se ajusta ya desde el momento de la puesta poniendo tantos huevos como pollos se van a criar (especies ajustadoras del tamaño de puesta), o (2) poniendo huevos de más y, posteriormente, ajustando el número de pollos según el alimento disponible (especies ajustadoras del número de pollos) (Lack, 1947). Esta clasificación no es estricta e inamovible, ya que la mayor parte de las especies ajustadoras del tamaño de puesta, en condiciones de escasez de alimento tienen la posibilidad de reducir el número de pollos (Soler, 2002). La estrategia del ajuste del número de pollos es seguida por aves como ardeidos, rapaces y la mayoría de los córvidos. Se basa en la eclosión asincrónica (ver apartado “incubación”) y consiste en que los pollos, al eclosionar de forma escalonada, mantienen una jerarquía de tamaño en el nido que provoca una competición intensa entre los pollos del nido por el alimento aportado por los padres. Esta competición conlleva la muerte por inanición de los pollos más pequeños cuando el alimento escasea (Slagsvold et al., 1995; Stenning, 1996). La estrategia ajustadora del número de pollos está considerada como una adaptación a condiciones en las que la disponibilidad de alimento es impredecible (Lack, 1947, 1966): se realiza un tamaño de puesta optimista y, cuando eclosionan los pollos, si no hay alimento suficiente para todos, muere el más pequeño, normalmente durante los primeros días de vida, cuando la inversión realizada en él es bastante pequeña. Para testar esta idea en una especie típicamente reductora del número de pollos como la Grajilla, Soler (1990) realizó un estudio en el que, tras calcular el número de huevos perdidos, pollos que mueren en cada clase de edad (de cinco en cinco días) y el número de pollos que vuelan, así como el peso medio y las Kilocalorías/gramo de cada una de las categorías mencionadas, encontró que, aunque los pollos que mueren suponen el 44,8% frente al 25,5% que llegan a volar (29,5% son los huevos que no llegan a eclosionar), la biomasa correspondiente a los pollos que mueren es el 38,4% frente al 58,1% que corresponde a los pollos volantones. Igualmente, un 31,6% de la energía producida corresponde a los pollos que mueren mientras que a los pollos volantones corresponde un 65,8%. Estos datos apoyan la idea de que la reducción de pollada es una adaptación: si las condiciones son muy buenas sobrevivirán todos los pollos, y, si las condiciones son malas, morirán los pollos más pequeños y durante los primeros días de edad, lo que supone un costo relativamente escaso tanto considerando la biomasa como considerando la energía producida. Heeb (1994) realizó un estudio experimental que apoyaba también esta idea, manipuló la asincronía en nidos de Grajilla y, en los nidos sincrónicos, murieron un mayor número de pollos que en los nidos asincrónicos.
Referencias Álvarez, F., Arias de Reyna, L. (1974). Reproducción de la urraca, Pica pica, en Doñana. Doñana, Acta Vert., 1: 77-96. Antikainen, E. (1978). The breeding adaptation of the Jackdaw Corvus monedula L. in Finland. Savonia, 2: 1-45. Arroyo, B. (1977). La avifauna de un cantil estepárico. Ardeola, 23: 41-47. Birkhead, T. R. (1991). The Magpies. T & A D Poyser, London. Coombs, F. (1978). The Crows. A Study of the corvids of Europe. Redwood Burn Limited Trowbeidge & Esher. London. Coulson, J. C. (1963). Egg size and shape in the Kittiwake (Rissa tridactyla) and their use in estimating the age and composition of populations. Proc. Zool. Soc. Lond. 140: 211-227. Cramp, S., Perrins, C. M. (1994). Handbook of the Bird of Europe the Middle East and North Africa. The Birds of the Western Palearctic. Volume VIII. Crows to Finches. Oxford University Press, Oxford. De Juana, E. (1980). Atlas ornitológico de La Rioja. Instituto de Estudios Riojanos, Logroño. Domínguez, L. (1999). Ecología de la Grajilla (Corvus monedula Linnaeus 1758) en la provincia de Madrid. Tesis doctoral. Univ. Complutense de Madrid. Madrid. Ejgelis, J. K. (1958). The biology of breeding of the Jackdaw (Coloeus monedula) in the oak forest “The wood on Worskla”. Vestnik Leningrad. Univ. 3, Biol., 1: 108-115. Elósegui, J. (1985). Navarra. Atlas de aves nidificantes. Caja de Ahorros de Navarra. Pamplona. Folk, C. (1968). Das insten und die Populationsdynamik der Dohle (Corvus monedula) in der CSSR. Zoologické Listy, 17: 221-236. Geroudet, P. (1955). Observaciones ornitológicas en la costa catalana. Ardeola, 2 (1): 31-56. Gibbons, D. W. (1987). Hatching asynchrony reduces parental investment in the Jackdaw. J. Anim. Ecol.,56: 403-414. Harrison, C. (1977). Guía de campo de los nidos, huevos y polluelos de las aves de España y de Europa (Norte de África y Próximo Oriente). Omega, Barcelona. Harrison, J. G. (1960). Notas sobre algunas aves de Andalucía (Mayo 1959). Ardeola, 6 (2): 340-343. Heeb, P. (1994). Intraclutch egg-mass var5iation and hatching asynchrony in the Jackdaw Corvus monedula. Ardea, 82: 287-297. Henderson, I. G., Hart, P. B. J. (1993). Provisioning, parental investment and reproductive success in Jackdaws Corvus monedula. Ornis Scand. 24: 142-148. Holyoak, D. (1967). Breeding biology of the Corvidae. Bird Study, 14: 153-168. Hoydt, D. F. (1979). Practical methods of estimating volume and fresh weight of eggs. Auk, 96: 73-77. Jonhsson, K. (1994). Colonial breeding and nest predation in the Jackdaw Corvus monedula using old Black Woodpecker Dryocopus martius holes. Ibis, 136: 313-317. Kirchner, H. (1933). Ueber das Brüten der Dohle in Kaminen. Beitr. Fortpfl. Biol. Bög., 9: 140. Kulczycki, A. (1973). Nesting of the members of the Corvidae in Poland. Acta Zool. Cracov., 13: 231-250. Lack, D. (1947). The significance of clutch size. Ibis, 89: 302-352. Lockie, J. D. (1955). The breeding and feeding of Jackdaws and Rooks with notes on Carrion Crows and other Corvidae. Ibis, 97: 341-369. Lorenz, K. (1931). Beiträge zur Ethologie sozialer Corvidae. J. F. Orn., 79: 67-127. Lorenz, K. (1982). Hablaba con las bestias, los peces y los pájaros. Labor S.A. Barcelona. Martin, T. E. (1995). Avian life-history evolution in relation to nest sites, nest predation and food. Ecol. Monogr. 65: 101-127. Mead, C. J., Peplar, G. R. M. (1975). Birds and other animals at Sand Martin colonies. Brit. Birds, 68: 89-99. Owen, J. H. (1930). Breeding habits of the Jackdaw. Brit. Birds, 24: 51-52. Owen, J. H. (1931). A note on the nesting of the Jackdaw. Brit. Birds, 24: 51-52. Quantz, B. (1930). Bemerkenswerte Dohlen-nistplätze. Beitr. Fortpfl. Biol. Vög., 6: 214. Ricklefs, R. E. (1967). A graphical method of fitting equations to growth curves. Ecology, 48: 978-983. Röell, A. (1978). Social behaviour of the Jackdaw, Corvus monedula, in relation to its niche. Behaviour, 64: 1-124. Rubio Argüello, M. C., Ena Álvarez, V. (1986). Fenología y productividad de la Grajilla (Corvus monedula) en la provincia de León. Alytes, 4: 105-114. Rubio-Arguello, C., Ena-Alvarez, V. (1987). Oología de la grajilla (Corvus monedula) en la Provincia de León (1984-1985). Pp. 209-217. En: Grupo Ibérico para el Estudio de la Avutarda (G.I.P.E.A.) (Eds.). I Congreso internacional de aves esteparias León (España) septiembre 1987. Dirección General de Montes, Caza, Pesca y Conservación de la Naturaleza y Consejería de Agricultura, Ganadería y Montes, León. Slagsvold, T., Amundsen, T., Dale, S. (1995). Costs and benefits of hatching asynchrony in blue tits Parus caeruleus. J. Anim. Ecol. 64: 563-578. Soler, J. J.; Møller, A. P., Soler, M. (1998). Nest building, sexual selection and parental investment. Evolutionary Ecology 12: 427-441. Soler, M. (1984). Biometría y biología de la Grajilla. Tesis doctoral. Universidad de Granada. Granada. Soler, M. (1987). Dimensiones, estructura, forma y disposición en la cavidad de los nidos de Corvus monedula. Doñana Acta Vertebrata, 14: 67-81. Soler, M. (1988a). Colour and shape of the eggshells of the jackdaw, Corvus monedula. A quantitative study. Gerfaut, 78: 383-396. Soler, M. (1988b). Egg size variation in the jackdaw Corvus monedula in Granada, Spain. Bird Study, 35: 69-76. Soler, M. (1988c). Nestling period variation in jackdaw Corvus monedula in relation to brood size. Bird Study, 36: 73-76. Soler, M. (1989a). Fracaso reproductor en Grajilla (Corvus monedula): pérdida de huevos y mortalidad de pollos. Ardeola, 36: 3-24. Soler, M. (1989b). Comienzo y duración de la incubación en grajilla (Corvus monedula L.). Studia Oecologica, 6: 387-394. Soler, M. (1990). Breeding success and productivity in the jackdaw (Corvus monedula L.) in Granada (Spain). Pp. 253-261. En: Pinowski, J., Summers-Smith, J. D. (Eds.). Granivorous birds in the agricultural landscape. Polish Scientific Publishers, Warsaw. Soler, M. (2002). Breeding strategy and begging intensity: influences on food delivery by parents and host selection by parasitic cuckoos. Pp. 413-427. En Wright, J., Leonard, M. L. (eds.) The Evolution of Begging. Kluwer Academic Publishers. Netherlands. Soler, M., Soler, J. J. (1987a). Selección del lugar de nidificación en Grajilla (Corvus monedula). Alytes, 5: 87-98. Soler, M., Soler, J. J. (1987b). Materiales utilizados por la Grajilla (Corvus monedula) en la construcción del nido. Alytes, 5: 99-120. Soler, M., Soler, J. J. (1987c). Fenología de puesta en Corvus monedula. Análisis de los factores que pueden afectarla. Ardeola, 34: 3-14. Soler, M., Soler, J. J. (1988). Egg size variation in the Jackdaw Corvus monedula in Granada, Spain. Bird Study, 35: 69-76. Soler, M., Soler, J. J. (1990). Crecimiento de los pollos de grajilla (Corvus monedula), relación entre desarrollo y comportamiento petitorio. Ardeola, 37 (1): 37-52. Soler, M., Soler, J. J. (1991b). Ritmo de actividad diaria y comportamiento en Grajilla (Corvus monedula) (Passeriformes, Corvidae) durante los períodos de construcción del nido y puesta-incubación. Zool. Baetica2: 35-45. Soler, M., Soler, J. J. (1991c). Clutch size variation in the Jackdaw, Corvus monedula, according to replacement clutches, breeding season and female age in southern Spain. Le Gerfaut, 81: 23-34. Soler, M., Soler, J. J. (1992a). Determinación de la distancia de recolección de los materiales blandos para el nido en la grajilla Corvus monedula. Etología, 2: 21-23. Soler, M., Soler, J. J. (1992b). Latitudinal trends in clutch size in single brooded hole nesting bird species: a new hypothesis. Ardea, 80: 293-300. Soler, M., Soler, J. J. (1993). Does the risk of nest predation affect clutch size in the Jackdaw Corvus monedula?. Bird Study 40: 232-235. Soler, M., Soler, J. J. (1996). Effects of experimental food provisioning on reproduction in the jackdaw Corvus monedula, a semi-colonial species. Ibis, 138: 377-383. Soler, M., Zúñiga, J. M., Camacho, I.(1982). Nidificación de Picus viridis en taludes de arcilla en ramblas de Guadix (Granada). Doñana, Acta Vert., 9: 195-209. Stenning, M. J. (1996). Hatching asynchrony, brood reduction and other rapidly reproducing hypothesis. Trends Ecol. Evol. 11: 243-246. Väisänen, R. A. (1969). Evolution of the Ringed Plover (Charadrius hiaticula L.) during the last hundred years in Europe, A new computer method based on egg dimensions. Ann. Acad. Sci. Fenn. A IV. 149: 1-90. Yom-Tov, Y. (1974). The effect of food and predation on breeding density and success, clutch size and laying date of the crow Corvus corone L.: an experimental approach. J. Anim. Ecol. 43: 479-498. Zimmermann, D. (1951). Zur Brutbiologie der Dohle, Coloeus monedula (L.). Orn. Beob., 48: 73-111.
Manuel Soler Fecha de publicación: 20-09-2006 Otras contribuciones: 1. Alfredo Salvador. 4-12-2014 Soler, M. (2016). Grajilla - Corvus monedula. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Morales, M. B. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||