|
|
|||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||
|
Para más información sobre cada apartado hacer click en:
|
Key words: Carrion crow, mate choice, cooperative breeding, nest, clutch size.
Biología de la reproducción Emparejamiento, comportamiento sexual y sistema reproductivo Emparejamiento: Las parejas reproductoras pueden formarse con individuos procedentes de un bando no reproductor y que se independizaron para buscar un territorio propio. También pueden ocurrir emparejamientos mediante reemplazo de uno de los individuos de la pareja territorial que ha muerto, por un inmigrante o un ayudante que ya formaba parte del grupo familiar (ver Formación del grupo más adelante). Las pautas de selección de pareja, cortejo y cópula de las cornejas españolas no han sido descritas todavía. Asumiendo que en España no difieren con respecto a los observados en otros lugares, a continuación se describen estos comportamientos registrados en el Reino Unido (Goodwin, 1986). - En el Reino Unido se ha detectado un vuelo poco frecuente pero con probable significación sexual así como territorial. Éste consiste en un vuelo con un deliberado batir de alas en el que las puntas de las mismas forman un arco más amplio que en vuelo normal (Goodwin, 1986). - Comportamiento de reverencia. Ambos sexos muestran comportamiento con carácter sexual en el que cada reverencia va seguida de una sacudida hacia arriba de la cabeza, cierre de las membranas nictitantes y despliegue de la cola (Goodwin, 1986). - Comportamiento de exhibición precopulatoria. El macho muestra el plumaje del cuerpo completamente despeinado, alas algo caídas y cola parcialmente desplegada mientras la hembra realiza un “despliegue de solicitud” en el que se agacha con las alas parcialmente abiertas y caídas y la cola temblorosa. (Goodwin, 1986). Apareamiento: El apareamiento se produce generalmente en el nido o cerca del mismo en algún árbol o en el suelo. Dura unos segundos y el macho aletea mientras mantiene el equilibrio sobre la espalda de la hembra. En parejas establecidas puede darse sin una exhibición previa conspicua (Goodwin, 1986). La cópula ocurre sólo justo antes del inicio de la puesta o durante la misma, a excepción de cópulas promiscuas con otros machos del grupo social. Sistema reproductor: La corneja se ha considerado durante mucho tiempo una especie socialmente monógama. Sin embargo estudios recientes en poblaciones cooperativas han demostrado que el sistema reproductor puede ser mucho más complejo. Individuos inmigrantes que forman parte del grupo social (sobre todo machos), con frecuencia se aparean con la hembra reproductora obteniendo descendencia que es criada junto a la de la pareja dominante (Baglione et al., 2002c). Por lo tanto, en las poblaciones de cornejas cooperativas existe también reproducción poliándrica. Formación del grupo social cooperativo En Europa el sistema social de la corneja ha sido invariablemente descrito como no cooperativo, es decir, sólo el macho y la hembra reproductores cuidan de su progenie (Tabla 1). Sin embargo, en una población urbana, en Suiza, Richner (1990) encontró un tercer cuidador (“ayudante”) en 2 de 33 nidos. En ambos casos el ayudante fue un hijo de esa pareja nacido 2 años antes que no se había dispersado. En el norte de España las cornejas se reproducen de forma cooperativa en el 73,3% de los territorios (Baglione et al., 2002a). En el sur de España, el 66,1% de los intentos de reproducción presentan cría cooperativa, observándose que la puesta tiene lugar antes que en parejas reproductoras (Roldán et al., 2013)1.
Tabla 1. Variación geográfica europea en la frecuencia de cría cooperativa de la corneja negra según Baglione et al. (2002a). 1 – Porcentaje de territorios de cría con presencia de individuos ayudantes
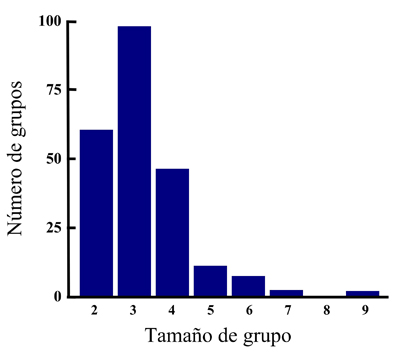
Aquí podemos encontrar cuatro tipos de grupos sociales: parejas reproductoras sin ayudantes, parejas con hijos de primer o segundo año (o mayores en raros casos) que no se han dispersado, parejas ayudadas por inmigrantes (sobre todo machos con cierto grado de parentesco con el macho reproductor del territorio, ver Baglione et al., 2003) y parejas ayudadas por descendencia no dispersada y también por inmigrantes. Los grupos sociales pueden estar formados por hasta 9 individuos (Figura 1, Baglione et al., 2002a).
Figura 1. Distribución del tamaño de grupo reproductor basado en cinco años de observaciones de 95 territorios en el norte de España (Baglione et al., 2002a).
La mayoría de los ayudantes de la pareja reproductora son juveniles del año anterior (67%), pero también se han observado individuos adultos. En Baglione et al. (2002b) se muestra experimentalmente cómo este comportamiento cooperativo no es de origen genético, sino dependiente de las condiciones ambientales. Esto se demostró trasfiriendo huevos de cornejas no cooperativas, de Suiza, a nidos pertenecientes a grupos cooperativos en el norte de España. Cinco de los seis pollos que llegaron a juveniles retrasaron su dispersión y dos de ellos mostraron comportamiento cooperativo en las siguientes temporadas de cría. Construcción del nido La construcción del nido empieza unos 15-20 días antes de la puesta. Normalmente en árboles altos situados en los límites de masas arbóreas, en pequeños parches o incluso aislados. El nido suele estar en el tercio superior del árbol, en una horquilla o rama cercana al tronco, según se ha descrito fuera de España (Cramp y Perrins, 1994). En el norte de España se ha estimado la densidad de nidos de corneja negra en 2 nidos / km2 (rango: 1.8 – 2.1) en varios años de observaciones de la misma zona (Baglione et al., 2002a). El transporte del material y la construcción del nido son llevados a cabo principalmente por los individuos reproductores, que también reciben ayuda de los demás miembros en el grupo familiar, si los hay. El tamaño del nido no se ha descrito en España. En otros lugares (Cramp y Perrins, 1994; Soler et al., 1998) el nido es grande aunque puede variar dependiendo de su localización: diámetro exterior: 30-47 cm, diámetro interior: 20 cm, altura: 30 cm, profundidad: 20 cm. La estructura no se ha descrito en España, pero en otros lugares suele constar de 4 capas: base de fuertes y cortas ramitas, a veces acompañadas con hojas que se mantienen unidas por una capa de musgo y hierba. A continuación ramitas más finas, tallos y raíces y, finalmente, un revestimiento de trozos de corteza, hierba, lana, plumas y material suave de origen antrópico. Puesta, huevos e incubación Las primeras puestas en Castilla y León se producen a mediados de marzo, a un ritmo de un huevo por día. Si la primera puesta falla, construirán un nido nuevo para realizar una puesta de reposición, incluso después de un segundo fracaso (muy raramente después de un tercero) si no está muy avanzada la temporada. La presencia de ayudantes aumenta la probabilidad de reposición después de un fracaso (Canestrari et al., 2008a). El tamaño medio de la primera puesta es de 4,6 huevos (rango 1-7, n = 588) y disminuye considerablemente en las puestas siguientes (ej. segunda puesta: 3,6 huevos, rango 1-6, n = 200); para más detalles consultar Canestrari et al. (2008a). En el sur de España los reproductores tempranos tienen un mayor tamaño de puesta y producen un número mayor de volanderos. Las puestas de reemplazo son muy raras y siempre sin éxito (Roldán et al., 2013)1. Los huevos son de forma sub-elíptica a oval, suaves y ligeramente brillantes. El color es azul a verde claro con patrones muy variables de manchas, motas, puntos y trazos de color verde oliva a marrón oscuro, a veces muy dispersas, otras veces oscureciendo el color de fondo, a menudo concentradas en un extremo; con gran variación dentro de la misma puesta. El tamaño medio del huevo no se ha descrito en España. En otros lugares es de 42,9 x 29,9mm, n = 500. Peso calculado: 19,8 g (Cramp y Perrins, 1994). La coloración de los huevos de corneja negra ha sido descrita a partir de colecciones de nidos conservadas en museos de historia natural por Soler et al. (2003). En la Tabla 2 se indican los valores medios de reflectancia en el espectro del ultravioleta, azul, verde, amarillo y rojo. Veintiséis huevos de corneja fueron medidos en siete nidos que tenían como característica haber sido parasitados por el críalo europeo (Clamator glandarius).
Tabla 2. Longitudes de onda (reflectancia) medidas en huevos de corneja negra por Soler et al. (2003). Las cifras indican la media + ES de siete nidos y están aproximadas a partir de la Figura 2 de Soler et al. (2003).
La incubación de los huevos es llevada a cabo tan sólo por la hembra reproductora durante unos 19-20 días. Durante este periodo la hembra es alimentada por el macho y otros miembros del grupo familiar si los hay, aunque a veces también abandona el nido para alimentarse por su cuenta. Cuidado de los pollos En las poblaciones cooperativas, no todos los miembros del grupo participan en el cuidado de los pollos ya que se ha visto que en aquellos con más de tres individuos, algunos pájaros rehúsan visitar el nido (Baglione et al., 2002a). Durante la primeras dos semanas la hembra se queda empollando en el nido y es alimentada por otros miembros del grupo. Cuando los pollos son pequeños, ella es la que los alimenta redistribuyendo la comida que el resto del grupo lleva al nido. Después, la hembra permite que los otros pájaros alimenten directamente a los pollos. La contribución individual al cuidado de los pollos en el nido varía de forma importante entre los miembros del grupo social. Los reproductores aportan más que los ayudantes y entre estos últimos los machos colaboran más que las hembras (Canestrari et al., 2005). La crianza de los pollos, su defensa, alimentación y cuidado, tienen unos costes energéticos importantes, de hecho, las cornejas pierden de media un 5% de su masa corporal durante la cría. Con experimentos de suministro de comida, Canestrari et al. (2007) demostraron que las cornejas son muy sensibles a dichos costes y que en condiciones favorables, de comida abundante durante la temporada de cría, prefieren reducir la pérdida de peso en lugar de invertir más en el cuidado de los pollos. Sin embargo los ayudantes, pero no los reproductores, sí pueden llegar a aumentar su ritmo de ceba de los pollos cuando el territorio que ocupan es de muy buena calidad y mantiene un elevado nivel de recursos tróficos a lo largo de todo el año (Canestrari et al., 2008).En grupos de cría cooperativa, el esfuerzo de aprovisionamiento de alimento varía entre los miembros del grupo, desde los que trabajan tanto como los reproductores hasta los que escasamente aportan alimento. Sin embargo, se ha demostrado experimentalmente que aquellos miembros de grupo que aportan poco incrementan su esfuerzo ante la falta de otro miembro (Baglione et al., 2010) 1. Canestrari et al. (2004) mostraron además, que los cuidadores evalúan el estado de hambre de los pollos cuando llegan al nido con comida y deciden si dársela toda, sólo una parte o no darles nada y alimentarse ellos. Estas “falsas cebas” ocurren en el 16,3% de las visitas, aunque el porcentaje varía mucho entre individuos, entre los que las hembras reproductoras poseen las tasas más altas. De esta forma, los cuidadores ajustan su gasto energético a las necesidades de los pollos. Se ha comprobado en grupos de cría cooperativa que las cornejas reproductoras incrementan la aportación falsa cuando experimentalmente se proporciona alimento suplementario a los pollos, mientras que los ayudantes no alteran su comportamiento; estos resultados sugieren que hay un compromiso entre las necesidades de los pollos y la condición de los que aportan alimento que modula la aportación falsa (Canestrari et al., 2010) 1. Supervivencia, periodo de volantón y madurez Tanto el éxito reproductor del grupo como la supervivencia de los pollos es muy variable según grupos y años. A continuación se muestran algunos de los resultados obtenidos en el estudio de una población del norte de la península a lo largo de 10 años (Canestrari et al., 2007): - Probabilidad de llegar a la eclosión de los huevos por territorio: Muy variable dependiendo del número de huevos puestos: 15 – 86%, n = 99 territorios. - Proporción de eclosión (excluyendo los nidos depredados): 79.9%, n = 99 territorios. - Volantones producidos por territorio exitoso: 0.65 – 1.24, n = 84 territorios. - Proporción de territorios que se reprodujeron con éxito por temporada (nidos donde se obtuvo al menos un volantón): 26.5 – 55.0%, n = 99 territorios. La presencia de ayudantes aumentó de forma significativa el éxito reproductor. Se considera a los pollos “volantones” a los 30 días, sin embargo, siguen dependiendo de los padres hasta 4-5 semanas después de dejar el nido. La precipitación media durante el periodo de nidificación se correlaciona positivamente con el número de volanderos (Roldán et al., 2013)1.
Demografía y dinámica de poblaciones En España, la madurez sexual la alcanzan a los dos años, aunque normalmente las hembras se reproducen por primera vez a los tres años. En otros lugares también se estima que las cornejas se reproducen a los tres años de edad (Moller, 2006). La tasa de supervivencia de las cornejas en España se estima en el 0.75 en los adultos (Baglione et al., 2005). Es una estima similar a la propuesta en una reciente revisión de varios parámetros demográficos de la corneja negra, según la cual se asigna una tasa de supervivencia para individuos adultos de corneja negra del 0.74 en base a 12.359 recuperaciones en el conjunto de Europa (Moller, 2006). Se desconoce la longevidad media en España, aunque en otros lugares se ha estimado en 19.0 años (Moller, 2006). No hay otros datos disponibles sobre la demografía y dinámica de poblaciones. Fuera del periodo reproductor se desconoce la proporción de sexos en España, la estructura de edades de la población y la supervivencia diferencial entre sexos y por edades. En el periodo reproductor la proporción de sexos se ha estimado en 1: 0.67 (77 individuos en 20 grupos reproductores, Baglione et al., 2002a) Las cornejas que se reproducen en grupo cooperativo ajustan la proporción de sexos de su descendencia según la composición del grupo. En el norte de España los hijos son más filopátricos y ayudan más en el nido que las hijas, que se dispersan antes y tienen mayores probabilidades de encontrar un sitio de reproducción. Las cornejas producen más hijos en grupos escasos en machos subordinados que en grupos con suficientes ayudantes, en los que se producen más hijas. Las hembras pueden desviar la sex-ratio primaria, consiguiendo la máxima probabilidad de vuelo a los hijos en el primer intento de reproducción y a las hijas en los siguientes. Este patrón se puede explicar por el mayor coste de producir machos que hembras, desviando las hembras hacia la producción de hijas en respuesta a los costes de los intentos de reproducción anteriores (Canestrari et al., 2012)1.
Referencias Baglione, V., Marcos, J. M., Canestrari, D. (2002a). Cooperatively breeding groups of carrion crow (Corvus corone corone) in northern Spain. Auk, 119: 790-799. Baglione, V., Canestrari, D., Marcos, J. M., Griesser, M., Ekman, J. (2002b). History, environment and social behaviour: experimentally induced cooperative breeding in the carrion crow. Proceeding of the Royal Society of London, Series B, 269: 1247–1251. Baglione, V., Canestrari, D., Chiarati, E., Vera, R., Marcos, J. (2010). Lazy group members are substitute helpers in carrion crows. Proceedings of the Royal Society B-Biological Sciences, 277 (1698): 3275-3282. Baglione, V., Canestrari, D., Marcos, J. M., Ekman, J. (2003). Kin selection in cooperative alliances of carrion crows. Science, 300: 1947-1948. Baglione, V., Marcos, J. M., Canestrari, D., Ekman, J. (2002c). Direct fitness benefits of group living in a complex cooperative society of carrion crows Corvus corone corone. Animal Behaviour, 64: 887–893. Baglione, V., Marcos, J. M., Canestrari, D., Griesser, M., Andreotti, G., Bardini, C., Bogliani, G. (2005). Does year-round territoriality rather than habitat saturation explain delayed natal dispersal and cooperative breeding in the carrion crow? Journal of Animal Ecology, 74: 842-851. Canestrari, D., Chiarati, E., Marcos, J. M., Ekman, J., Baglione, V. (2008b). Helpers but not breeders adjust provisioning effort to year-round territory resource availability in carrion crows. Animal behaviour, 76: 943-949. Canestrari, D., Marcos, J. M., Baglione, V. (2004). False feedings at the nest of carrion crows Corvus corone corone. Behavioural Ecology and Sociobiology, 55: 477-483. Canestrari, D., Marcos, J. M., Baglione, V. (2005). Effect of parentage and relatedness on the individual contribution to cooperative chick care in carrion crows. Behavioral Ecology and Sociobiology, 52: 422-428. Canestrari, D., Marcos, J. M., Baglione, V. (2007). Costs of chick provisioning in cooperatively breeding carrion crows: an experimental study. Animal Behaviour, 73: 349-357. Canestrari, D., Marcos, J. M., Baglione, V. (2008a). Reproductive success increases with group size in cooperative Carrion Crows Corvus corone corone. Animal Behaviour, 75: 403-416. Canestrari, D., Vera, R., Chiarati, E., Marcos, J. M., Vila, M., Baglione, V. (2010). False feeding: the trade-off between chick hunger and caregivers needs in cooperative crows. Behavioral Ecology, 21 (2): 233-241. Canestrari, D., Vila, M., Marcos, J. M., Baglione, V. (2012). Cooperatively breeding carrion crows adjust offspring sex ratio according to group composition. Behavioral Ecology and Sociobiology, 66 (9): 1225-1235. Charles, J. K. (1972). Territorial behavior and the limitation of population size in Crows Corvus corone and Corvus cornix. Ph.D. dissertation, University of Aberdeen. Richner, H. (1990). Helpers at the nest in Carrion Crows Corvus corone corone. Ibis, 132: 105–108. Cramp, S., Perrins, C. M. (1994). Handbook of the Bird of Europe the Middle East and North Africa. The Birds of the Western Palearctic. Volume VIII. Crows to Finches. Oxford University Press, Oxford. Goodwin, D. (1986). Crows of the world. Natural History Museum Publications, London. Moller, A. P. (2006). Sociality, age at first reproduction and senescence: comparative analyses of birds. Journal of evolutionary biology, 19: 682-689. Richner, H. (1990). Helpers at the nest in Carrion Crows Corvus corone corone. Ibis, 132: 105–108. Roldán, M., Martín-Gálvez, D., Rodríguez, J., Soler, M. (2013). Breeding biology and fledgling survival in a Carrion Crow Corvus corone population of southern Spain: a comparison of group and pair breeder. Acta Ornithologica, 48 (2): 221-235. Soler, J. J., Avilés, J. M., Soler, M., Møller, A. P. (2003). Evolution of host egg mimicry in a brood parasite, the great spotted cuckoo. Biological Journal of the Linnean Society, 79: 551–563. Soler, J. J., Moller, A. P., Soler, M. (1998). Nest building, sexual selection and parental investment. Evolutionary Ecology, 12: 427-441. Wittenberg, J. (1988). Langfristige Entwicklung einer Population der Rabenkrähe (Corvus c. corone) bei Braunschweig, ihre Zusammensetzung und ihr Einfluss auf andere Arten. Beiheft Veröffentlichungen Naturschutz und Landschaftspflege Baden-Württemberg, 53: 2111–2223.
Diana Bolopo1, Daniela Canestrari2, Vittorio Baglione1,3 1Dpto. Ciencias Agroforestales (Área de Zoología), Escuela Técnica Superior de Ingenierías Agrarias, Universidad de Valladolid, 34004 Palencia 2 Dpto. de Biología Animal, Universidad de Granada, Campus de Fuente Nueva, 18071 Granada 3 Instituto Universitario de Gestión Forestal Sostenible UVA-INIA, Valladolid Fecha de publicación: 10-11-2009 Otras contribuciones: 1. Alfredo Salvador. 22-04-2015 Bolopo, D., Canestrari, D., Baglione, V. (2015). Corneja negra – Corvus corone. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Morales, M. B. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||