|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: Cory's Shearwater, movements, migration, dispersal.
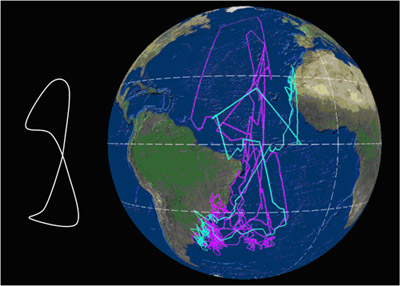
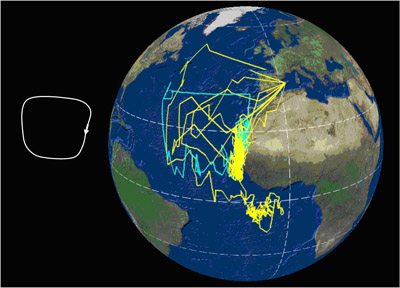
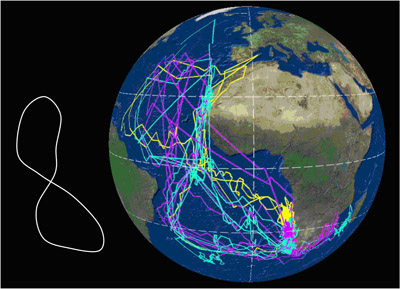
Movimientos Migración Se trata de una especie que realiza grandes migraciones, buena parte de las cuales son transecuatoriales. Una vez finalizado el período reproductor, entre mediados y finales de octubre, los individuos inician la migración postnupcial, que los mantendrá alejados de las áreas de cría unos 100 días en promedio, de los cuales pasarán 80 en las zonas de invernada y el resto en el trayecto migratorio. La migración prenupcial, de retorno a las colonias de cría, tiene lugar mayoritariamente durante febrero, terminando el viaje a lo largo de febrero y marzo (Reyes-González et al., 2017). Las pardelas cenicientas atlánticas de las poblaciones españolas migran hacia diferentes áreas de invernada. Las costas de Namibia y corriente de Benguela representan la principal área (40,5%), seguido de la corriente de Agujas (21,5%), la confluencia de las corrientes de Agujas y Benguela (13,5%), corriente de Canarias (11,5%), aguas de Brasil (10%) y aguas del Atlántico sur central (3%). Ningún ejemplar de colonias españolas, en 6 años de estudio, visitó el noroeste del Atlántico para invernar (Reyes-González et al., 2017). Sin embargo, en la población de Salvajes un 4% de los animales utiliza también esta última zona (Dias et al, 2012). Durante la migración hacen 1-2 paradas que duran en promedio siete días (rango= 1-31 días). Las aves que no hacen detenciones migran más rápido (media= 535 km/día-1), que las que hacen detenciones (media= 428 km/día-1). Las aves que abandonan más tarde las colonias emigran más rápido que las que lo hacen antes. En total, la velocidad media a la ida fue 429 km/día-1 y 644 km/día-1 a la vuelta. Cuando están en migración, emplean el 50% del tiempo volando y lo hacen mucho más durante el día que durante la noche. Las fases lunares afectan a la actividad migratoria, volando más en noches de luna llena (Dias et al., 2012)1. El sentido del olfato juega un importante papel en la orientación durante la navegación. Ejemplares de pardela cenicienta atlántica que fueron desprovistos temporalmente de su capacidad olfativa y desplazadas a 800 km de su colonia de cría en las islas Azores, donde la actividad es exclusivamente nocturna, fueron incapaces de volver allí (Gagliardo et al., 2013)1; en cambio, ejemplares de las islas Salvajes, población con actividad diurna y nocturna, en un experimento similar no pudieron volver de noche teniendo que esperar a hacerlo de día (Dell'Ariccia y Bonadonna, 2013)1. La fidelidad a las áreas de invernada es muy elevada. En poblaciones españolas, cerca del 77% de los animales, seguidos a lo largo de hasta 6 años, tuvieron una única área de invernada. Tan sólo un 22% presentó dos áreas de invernada, y poco más del 1% invernó en tres áreas diferentes. En promedio, a nivel individual los animales presentaron siempre un solapamiento espacial mayor del 60% entre años en sus zonas de invernada (Reyes-González et al., 2017). En ejemplares de las Islas Salvajes, se ha encontrado consistencia individual entre años en la ruta migratoria en la primera parte del viaje hacia el sur, entre 10ºN y 20ºS, pero no en el resto ni en la vuelta (Dias et al., 20131). La especie presenta migración parcial, dado que un pequeño número de individuos permanece cada invierno en el área cercana a la colonia de cría. En las poblaciones españolas, un 11,5% de los animales (n=237) que se reproducen en las Islas Canarias permanece en la corriente de Canarias durante los meses de invierno (Reyes-González et al., 2017). En la población de las islas Salvajes, un 8,1% de los individuos (n= 172) monitoreados permanecieron en aguas cercanas a la colonia, en el entorno de la Corriente de Canarias (Pérez et al., 2014). Casi todos eran machos y en años diferentes variaron su estrategia entre permanecer cerca o migrar al hemisferio sur. Los machos que se quedaron cerca ocuparon antes el nido que los que migraron, lo que podría ser ventajoso (Pérez et al., 2014) 1. La respuesta de estrés, medida en niveles de corticosterona en plumas, varía entre sexos, años y estrategias migratorias. Fueron más elevados en machos que en hembras y mayores en machos que migraron a áreas de invernada alejadas de la colonia en comparación con los que permanecieron cerca. Además, las aves que acumularon niveles más altos de corticosterona en invierno tuvieron más éxito reproductivo (Pérez et al., 2016)1. Se ha demostrado experimentalmente que el nivel de inversión parental en una temporada e reproducción condiciona la estrategia de migración posterior y su calendario, con efectos que se extienden a la siguiente temporada de reproducción. Las aves con menor inversión en la reproducción, es decir, aquellas que fracasan en la reproducción (en el caso experimental, aquellas a las que se privó del pollo recién nacido), iniciaron antes tanto la migración postnupcial como la migración prenupcial, retornando antes a la colonia de cría en la primavera siguiente. Los machos manipulados migraron en mayor proporción a áreas más cercanas, lo que sugiere que la migración parcial es dependiente de la condición física. La probabilidad de nidificación al año siguiente fue mayor en las aves manipuladas (es decir, las que fracasaron en la cría) que en las aves control (Catry et al., 2013)1. El viaje completo desde las áreas de cría a las de invernada y retorno puede abarcar entre 15.000 y 35.000 km, y puede seguir varias estrategias (Figura 1). En esencia, las trayectorias de las aves describen un lazo a lo largo del Atlántico. Algunas áreas de invernada son compartidas con C. diomedea, aunque en proporciones claramente distintas (González-Solís et al., 2007). Los factores determinantes de las rutas de migración aún no están claros, aunque el viento parece ser el factor más influyente en la elección de la ruta y el calendario migratorio, pues las pardelas aprovechan los vientos dominantes para disminuir el coste energético a pesar de conllevar rutas más largas, vientos que además aparecen en determinados momentos del año (Felicísimo et al., 2008; González-Solís et al., 2009). Ello explica la ruta migratoria en forma de lazo con la que C. borealis cruzan dos veces el Atlántico para llegar a las corrientes de Benguela y Agulhas.
Figura 1. Cada línea corresponde a la migración anual de una pardela; los colores distinguen las poblaciones de origen (amarillo= C. diomedea de las islas Baleares, azul = C. borealis de las Islas Canarias; magenta = C. borealis de Islas Azores). Datos obtenidos mediante el uso de geolocalizadores por niveles de luz; los puntos sobre tierra firme corresponden a errores de localización intrínsecos al método. Arriba: Rutas seguidas por pardelas reproductoras en Canarias (1) y Azores (4) que pasaron el invierno en aguas de la plataforma continental de América del Sur, en aguas de la Corriente de Brasil; Centro: rutas seguidas por 5 pardelas reproductoras en las Canarias (1) y Baleares (4) para pasar el invierno boreal en aguas de la Corriente Canaria; Abajo: rutas migratorias seguidas por 11 pardelas cenicientas reproductoras en las Islas Canarias (4), Azores (5) y Baleares (2), con su principal cuartel de invernada en el área de la Corriente de Benguela y la Corriente de Agulhas (Océano Índico). Según González-Solís et al. (2009).
Dispersión A pesar de su carácter altamente filopátrico, su gran capacidad migratoria ligada a la congregación de individuos de diverso origen en los mismos cuarteles de invernada (González-Solís et al., 2007) apuntan a un pequeño flujo génico entre poblaciones, seguramente debido a que, al migrar normalmente en grupos, algunos ejemplares podrían seguir a efectivos pertenecientes a otras colonias reproductoras. La proporción de machos que se reprodujeron por primera vez en la isla Selvagem Grande y que habían nacido allí varió entre años del 42,4% al 63,9% (Mougin et al., 2000c)1. Como prueba de intercambio entre pardela atlántica y pardela cenicienta hay numerosos casos reportados de aves de C. borealis reproduciéndose en colonias de C. diomedea (Lo Valvo y Massa, 1988; Granadeiro, 1991; Sánchez, 1997; Thibault y Bretagnolle, 1998; Gómez-Díaz et al., 2009). Se ha estimado mediante análisis genéticos que por cada generación habrían migrado 90 individuos de C. borealis a C. diomedea y 55 individuos desde C. diomedea a C. borealis (Genovart et al., 2013)1.
Referencias Catry, P., Dias, M. P., Phillips, R. A., Granadeiro, J. P. (2013). Carry-over effects from breeding modulate the annual cycle of a long-distance migrant: an experimental demonstration. Ecology, 94 (6): 1230-1235. Dell'Ariccia, G., Bonadonna, F. (2013). Back home at night or out until morning? Nycthemeral variations in homing of anosmic Cory's shearwaters in a diurnal colony. Journal of Experimental Biology, 216 (8): 1430-1433. Dias, M. P., Granadeiro, J. P., Catry, P. (2012). Do Seabirds Differ from Other Migrants in Their Travel Arrangements? On Route Strategies of Cory's Shearwater during Its Trans-Equatorial Journey. Plos One, 7 (11): e49376. Dias, M. P., Granadeiro, J. P., Catry, P. (2013). Individual variability in the migratory path and stopovers of a long-distance pelagic migrant. Animal Behaviour, 86 (2): 359-364. Dias, M. P., Granadeiro, J. P., Phillips, R. A., Alonso, H., Catry, P. (2010). Breaking the routine: individual Cory's shearwaters shift winter destinations between hemispheres and across ocean basins. Proceedings of the Royal Society B: Biological Sciences, 278: 1786-1793. Felicísimo, A. M., Muñoz, J., González-Solís, J. (2008). Ocean surface winds drive dynamics of transoceanic aerial movements. Plos One, 3: e2928. Gagliardo, A., Bried, J., Lambardi, P., Luschi, P., Wikelski, M., Bonadonna, F. (2013). Oceanic navigation in Cory's shearwaters: evidence for a crucial role of olfactory cues for homing after displacement. Journal of Experimental Biology, 216 (15): 2798-2805. Genovart, M., Thibault, J. C., Igual, J. M., Bauza-Ribot, M. M., Rabouam, C., Bretagnolle, V. (2013). Population Structure and Dispersal Patterns within and between Atlantic and Mediterranean Populations of a Large-Range Pelagic Seabird. Plos One, 8 (8): e70711. Gómez-Díaz, E., González-Solís, J., Peinado, M. A. (2009). Population structure in a highly pelagic seabird, the Cory's shearwater Calonectris diomedea: an examination of genetics, morphology and ecology. Marine Ecology Progress Series, 382: 197-209. González-Solís, J., Croxall, J. P., Oro, D., Ruiz, X. (2007). Trans-equatorial migration and mixing in the wintering areas of a pelagic seabird. Frontiers in Ecology and the Environment, 5: 297-301. González-Solís, J., Felicisimo, A., Fox, J. W., Afanasyev, V., Kolbeinsson, Y., Muñoz, J. (2009). Influence of sea surface winds on shearwater migration detours. Marine Ecology Progress Series, 391: 221-230. Granadeiro, J. P. (1991). On Cory's shearwater ringed at Selvagem Grande, Madeira (30º09´N, 15º52´W) and recovered on Berlenga island, Portugal (39º24´N, 93º0´W). Bocagiana, 145: 1-4. Igual, J. M., Afán, I. (2005). Pardela cenicienta. En: Molina, B., Lorenzo, J. A., López-Jurado, C. (eds.). Noticiario Ornitológico. Ardeola, 52: 423-435. Lo Valvo, M., Massa, B. (1988). Considerations on a specimen of Cory´s shearwater ringed at Selvagem Grande and recovered in the central Mediterranean. Bocagiana, 124: 1-5. Mougin, J. L., Jouanin, C., Roux, F. (2000c). Démographie du puffin cendré Calonectris diomedea de Selvagem Grande. Rev. Ecol. (Terre Vie), 55: 275-290. Pérez, C., Granadeiro, J. P., Dias, M. P., Alonso, H., Catry, P. (2014). When males are more inclined to stay at home: insights into the partial migration of a pelagic seabird provided by geolocators and isotopes. Behavioral Ecology, 25 (2): 313-319. Pérez, C., Granadeiro, J. P., Dias, M. P., Catry, P. (2016). Sex and migratory strategy influence corticosterone levels in winter-grown feathers, with positive breeding effects in a migratory pelagic seabird. Oecologia, 181 (4): 1025-1033. Reyes-González, J. M., Zajková, Z., Morera Pujol, V., de Felipe, F., Militão, T., Dell’Ariccia, G., Ramos, R., Igual, J. M., Arcos, J. M., González-Solís, J. (2017). Migración y ecología espacial de las poblaciones españolas de pardela cenicienta. Monografía n.º 2 del programa Migra. SEO/BirdLife, Madrid. Sánchez, A. (1997). La Pardela cenicienta (Calonectris diomedea) en las islas Columbretes: biología y conservación. Universidad de Valencia, Valencia. Thibault, J. C., Bretagnolle, V. (1998). A Mediterranean breeding colony of Cory´s shearwater Calonectris diomedea in which individuals show behavioural and biometric characters of the Atlantic subspecies. Ibis, 140: 253-258.
José Manuel de los Reyes González y Jacob González-Solís Bou Fecha de publicación: 28-11-2016 Otras contribuciones: 1. Alfredo Salvador. 28-11-2016 Reyes-González, J. M., González-Solís, J. (2016). Pardela cenicienta atlántica - Calonectris borealis. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Morales, M. B. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|
|
|
|
|
|