|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: Fire Salamander, description, size, variation.
Sinónimos y combinaciones Lacerta salamandra Linnaeus, 1758; Salamandra candida Laurenti, 1768; Salamandra maculosa Laurenti, 1768; Salamandra terrestris Houttuyn, 1782; Gecko salamandra Meyer, 1795; Triton vulgaris Rafinesque, 1814; Salamandra maculata Merrem, 1820; Salamandra vulgaris Cloquet, 1827; Triton corthyphorus Leydig, 1867; Salamandra maculosa var. nera Doderlein, 1872; Salamandra maculosa var. pezzata Doderlein, 1872; Salamandra maculosa Boulenger, 1882; Salamandra maculosa var. europaea Bedriaga, 1883; Salamandra maculosa var. gallaica López-Seoane, 1885; Salamandra maculosa var. molleri Bedriaga, 1889; Salamandra salamandra Lönnberg, 1896; Salamandra maculosa var. nigriventris Dürigen, 1897; Salamandra maculosa var. typica Bedriaga, 1897; Salamandra maculosa var. taeniata Dürigen, 1897; Salamandra maculosa var. quadri-virgata Dürigen, 1897; Salamandra maculosa var. coccinea Schweizerbarth, 1909; Salamandra salamandra salamandra Poche, 1912; Salamandra moncheriana Schreiber, 1912; Salamandra maculata Schreiber, 1912; Salamandra maculosa forma fastuosa Schreiber, 1912; Salamandra maculosa var. speciosa Schreiber, 1912; Salamandra salamandra gallaica Nikolskii, 1918; Salamandra salamandra molleri Nikolskii, 1918; Salamandra salamandra taeniata Nikolskii, 1918; Salamandra maculosa taeniata forma bernardezi Wolterstorff, 1928; Salamandra salamandra carpathica Calinescu, 1931; Salamandra maculosa bejarae Wolterstorff, 1934; Salamandra maculosa bejarae Wolterstorff, 1934; Salamandra salamandra almanzoris Müller and Hellmich, 1935; Salamandra maculosa subsp. Bernardezi Scharlinski, 1939; Salamandra salamandra albanica Gayda, 1940; Salamandra salamandra thuringica Gayda, 1940; Salamandra salamandra bejarae Mertens and Müller, 1940; Salamandra salamandra hispanica Mertens and Müller, 1940; Salamandra salamandra bonnali Mertens and Müller, 1940; Salamandra salamandra bernardazi Mertens and Müller, 1940; Salamandra salamandra forma francica Sochurek and Gayda, 1941; Salamandra salamandra forma werneri Sochurek and Gayda, 1941; Salamandra salamandra gigliolii Eiselt and Lanza, 1956; Salamandra salamandra salamandra Eiselt, 1958; Salamandra salamandra fastuosa Eiselt, 1958; Salamandra salamandra terrestris Eiselt, 1958; Salamandra (Salamandra) salamandra Özeti, 1967; Salamandra salamandra beschkovi Obst, 1981; Salamandra salamandra (gallaica) crespoi Malkmus, 1983; Salamandra salamandra morenica Joger and Steinfartz, 1994; Salamandra salamandra longirostris Joger and Steinfartz, 1994; Salamandra salamandra alfredschmidti Köhler and Steinfartz, 2006 (Frost, 2014).
Origen y evolución La historia biogeográfica de S. salamandra es bastante compleja. Los datos genéticos analizados hasta la fecha apuntan a una colonización del centro y norte de Europa en el período postglacial (Steinfartz et al., 2000). Esta colonización debió originarse desde al menos dos áreas geográficas situadas en el sur de Europa: la Península Ibérica y la Península Balcánica (Steinfartz et al., 2000). Sin embargo, la enorme diversidad y diferenciación genética observada entre las subespecies de la Península Ibérica (Dopazo et al., 1998; García-París et al., 2003; Velo-Antón et al., 2007) indican un proceso de diversificación anterior a la colonización del centro y norte de Europa. Los diferentes ciclos climáticos ocurridos durante el Plioceno-Pleistoceno llevarían a esta especie a buscar micro-refugios dentro de la Península Ibérica, conduciendo a procesos de subespeciación alopátrica. Durante periodos con condiciones climáticas más propicias para esta especie, se producirían expansiones desde los microrefugios, dando lugar a las diferentes zonas de contacto existentes actualmente. Este escenario podría explicar la enorme complejidad morfológica y el polimorfismo observado en esta especie. Un ejemplo de la complejidad biogeográfica de S. salamandra es la agrupación, a nivel mitocondrial de las subespecies S. s. bernardezi (restringida al noroeste de la Península Ibérica) y S. s. gigliolii (distribuida por el sur de la Península Itálica) en un grupo monofilético (Steinfartz et al., 2000; García-París et al., 2003; Vences et al., 2014), el cual está claramente diferenciado del resto de subespecies. Nuevos datos genéticos, basados en secuencias de un gen nuclear, apoyan el estrecho parentesco entre estas dos subespecies y su diferenciación genética con el resto, a pesar de ser muy diferentes en caracteres morfológicos y en el patrón de coloración (Velo-Antón et al., 2015a). Ambos grupos podrían considerarse como poblaciones relictas de una gran población distribuida ancestralmente por Europa, que fue afectada por los diferentes ciclos climáticos durante el Pleistoceno (Steinfartz et al., 2000), existiendo evidencias de posteriores recolonizaciones más recientes de ambas subespecies, tanto en la Península Ibérica (hacia los Pirineos, García-París et al., 2003), como en la Península Itálica (hacia el norte de Italia, D. Canestrelli, comunicación personal). Por otro lado, la separación de S. s. longirostris, el linaje más diferenciado dentro de los presentes en la Península Ibérica, probablemente es anterior, quizás en el Plioceno o en el Pleistoceno temprano, y actualmente se mantiene aislada geográficamente con respecto al resto de subespecies (García-París et al., 1998). Es necesario un análisis ecológico y genético de las poblaciones del tercio sur de la Península Ibérica para determinar cuando ocurrió la separación entre las poblaciones del sur y analizar el flujo genético histórico y presente.
Descripción del adulto La descripción general de esta especie se realiza en base a la subespecie con mayor distribución (S. s. gallaica) y a continuación se describen las características morfológicas y rango de distribución para cada una de las subespecies descritas en la Península Ibérica. Aunque estas descripciones recogen las características generales de cada subespecie, hay que reiterar que, dada la complejidad de cada grupo, se observan grandes variaciones sobre los patrones generales descritos. La piel es lisa y brillante y el diseño de coloración, con manchas o líneas amarillas, o una combinación de ambas, sobre fondo negro, es muy variable entre subespecies, pero también dentro de cada subespecie. Es común la presencia de una mancha amarilla en la parte superior del inicio de cada extremidad, mancha ya visible durante las últimas fases del desarrollo larvario. En ejemplares de algunas subespecies (ver descripción de cada subespecie) el fondo puede ser también marrón oscuro y pueden presentar manchas de color rojo o vino. El vientre es generalmente negro aunque varía a nivel intra- e interpoblacional, y las glándulas parótidas suelen ser amarillas y muy prominentes. El tamaño y la forma de las manchas amarillas en el cuerpo se modifican con el tiempo, como ocurre en otras especies del género (Beukema, 2011), haciendo que el patrón de manchas no sea un sistema adecuado para el reconocimiento de individuos. Se han descrito ejemplares albinos (Benavides et al., 2000; Boada et al., 2012), así como casos de melanismo y neotenia (Palaus i Soler, 1999; Martínez y Fesser Justes, 2000; Salvador y García París, 2001) y otras coloraciones anómalas (Rivera et al., 1994). Se asume que el patrón de coloración general (negro y amarillo) cumple funciones aposemáticas (e.g., Llorente et al., 1995; Vences et al., 2014), aunque también se han discutido sus posibles funciones crípticas y miméticas (Bas y Gasser, 1994; Rivera et al., 2014). La cabeza está bien diferenciada del cuerpo. Es grande y robusta, con un morro puntiagudo o redondeado, carácter usado para diferenciar subespecies. Los ojos son grandes y de color negro, con el iris de color marrón oscuro. El tamaño y la forma de las glándulas parótidas son también muy variables y están cubiertas por poros glandulares. El cuerpo es casi cilíndrico, a excepción de la zona ventral, con surcos transversales a lo largo del cuerpo y ausencia de crestas caudales o dorsales. Presenta poros glandulares por todo el cuerpo, y en las líneas laterales del cuerpo se distribuyen en hileras paralelas. La cola es de aspecto cilíndrico y su diámetro disminuye en sentido dorso-caudal. El tamaño de las extremidades y los dedos, desprovistos de membranas digitales, es también muy variable (Barbadillo, 1987). Se han encontrado casos de polimelia en Málaga (Escoriza y García-Cardenete, 2006) y en Asturias (Villanueva, 2007). La hendidura cloacal es alargada y los labios cloacales son más abultados en los machos, especialmente durante la época de celo.
Dimorfismo sexual No presenta un dimorfismo sexual muy marcado, aunque las hembras son más robustas y alcanzan mayor longitud que los machos (Galán y Fernández-Arias, 1993; Cordero et al., 2007; Labus et al., 2013; Balogová y Uhrin 2015), tienen los dedos de todas las extremidades más robustos (González de la Vega, 1988), cabezas más grandes y las colas y extremidades más cortas (Labus et al., 2013). Los labios cloacales son de mayor tamaño en los machos, haciéndose más evidente esta diferencia durante la época nupcial. Recientemente se ha demostrado que existe dicromatismo sexual en poblaciones eslovacas (Balogová y Uhrin, 2015), observándose un mayor tamaño de las manchas amarillas en los machos, pero no en el número de manchas o en la tonalidad del color amarillo.
Tamaño Es un urodelo de tamaño medio y aspecto robusto que no suele superar los 200 mm de longitud total, aunque en alguna población llega a alcanzar los 250 mm (Ferrand de Almeida et al., 2001).
Descripción de los huevos y de la larva Huevo Los huevos de la salamandra común son blancos y con un diámetro aproximado de entre 4.5-4.7 mm, rodeados por una cápsula transparente (Buckley, 2003; Buckley et al., 2007; Galán et al., 2011). Larva Existen en las poblaciones ovovivíparas (larvíparas). El tamaño de la larva oscila entre 19 y 35 mm antes de la metamorfosis (Velo-Antón et al., 2015b), aunque se han observado larvas de hasta 70 mm en las lagunas de Gredos (Alcobendas y Castanet, 2000). La cabeza es robusta y ancha, y el morro muy redondeado. Presenta una cresta que comienza por el lado dorsal a mitad del cuerpo continuando a lo largo de la cola, y por el lado ventral iniciándose al comienzo de la cola, formando un extremo romo o redondeado al final de la cola. Las branquias están formadas por tres penachos branquiales con numerosas ramificaciones (Díaz-Paniagua y López-Jurado, 1981). El patrón de ramificación es bastante variable y está relacionado inversamente con la cantidad de oxígeno en el agua y el momento de desarrollo al ser liberadas al agua. Se observa una tendencia al aumento de tamaño y complejidad de las branquias con el descenso de la concentración de oxígeno en el agua (Campeny, 1991). Durante el desarrollo de la larva y la metamorfosis, las crestas dorsal y caudal van desapareciendo, y las branquias se van reabsorbiendo progresivamente. El color pardo y moteado de negro inicial va oscureciéndose y distinguiéndose manchas o líneas amarillas. Las manchas claras del inicio de las extremidades cambian al color amarillo y el vientre se vuelve grisáceo. Al final de la metamorfosis pueden alcanzar los 80 mm. Se ha encontrado casos de bicefalia en larvas de esta especie (Velo-Antón et al., 2007a).
Características citogenéticas y bioquímicas Salamandra salamandra posee 24 cromosomas, metacéntricos o submetacéntricos, exceptuando el par 8 que presenta carácter heterobraquial (Mancino et al., 1969). En las regiones proximales de los cromosomas muestra una granulación (Ragghianti et al., 1982), en lugar de una banda definida, diferenciándose así de especies como la salamandra rabilarga (Chioglossa lusitanica). El análisis bioquímico mediante proteínas de suero identificó dos series de proteínas de tipo seroalbúmina (Gasser, 1967) y una gran variabilidad en las fracciones séricas y hasta 22 globulinas diferentes (Gasser, 1978; Montori y Herrero, 2004). En S. salamandra el contenido en ADN por núcleo diploide es de 70,55 pg. (Lizana et al., 2000).
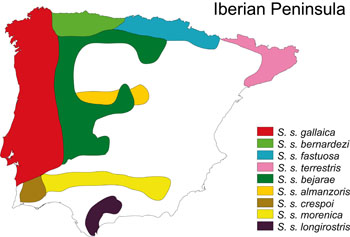
Variación geográfica Esta especie presenta una enorme variabilidad morfológica, morfométrica, de diseños de pigmentación y de formas de reproducción. Como resultado, se han descrito hasta 10 subespecies en la Península Ibérica (Figura 1). Algunas de estas variaciones no están restrictas a diferencias intersubespecíficas o interpoblaciones, sino que pueden ocurrir también dentro de las subespecies o poblaciones. Esta especie muestra politipismo pero también polimorfismo y se hace necesaria una revisión de la validez de la taxonomía subespecífica, tal y como se ha realizado recientemente con S. s. alfredschmidti (Köhler y Steinfartz, 2006), la cual constituye un ejemplo del polimorfismo de color y no de una diferenciación ecológica o genética con S. s. bernardezi (Beukema et al., 2015). Además, la existencia de zonas de contacto e introgresión entre algunas de estas subespecies donde se observa individuos con caracteres morfológicos intermedios (Hillenius, 1968; Fachbach, 1976; Bas, 1983; Malkmus, 1983; Bas y Gasser, 1994; Veith, 1997; Alcobendas et al., 1996; García-París et al., 2003; Galán, 2007; Reis et al., 2011), y la aparente invalidez de los caracteres morfométricos usados en la descripción de subespecies (Montori y Francesch, 1995), refuerzan el interés por una re-evaluación de la taxonomía dentro esta especie, la cual debería realizarse combinando estudios morfológicos, genéticos y ecólogicos. Estudios de sistemática molecular (Alcobendas et al., 1994; García-París et al., 1998, 2003; Buckley y Martínez-Solano, datos no publicados) recuperan cinco linajes evolutivos principales dentro de la especie, linajes definidos a partir de su variación en marcadores del ADN mitocondrial: el complejo bernardezi, de poblaciones vivíparas o de estrategia mixta; el complejo gallaica/bejarae/terrestris/fastuosa, al oeste y centro de la Península, extendiéndose también por el resto de Europa; la subespecie S. s. almanzoris, en torno al Sistema Central; el complejo crespoi/morenica, entre los ríos Guadiana y Guadalquivir, y la subespecie S. s. longirostris, localizada al sur del Guadalquivir. La complejidad dentro de estos grandes linajes es notable, en especial en los dos primeros. Además, los contactos secundarios entre estos linajes históricos y la introgresión genética entre ellos, como en el caso de las subespecies S. s. bernardezi y S. s. fastuosa, dificultan aún más el establecer una taxonomía intraespecífica robusta. A falta de un estudio concluyente que analice las diferencias morfológicas y genéticas entre las subespecies descritas en la península Ibérica, se describen a continuación las nueve subespecies (Figura 1) hasta ahora reconocidas (con la excepción de S. s. alfredschmidti, ver Beukema et al., 2015), enfatizando que su validez está todavía por clarificar.
Figura 1. Distribución aproximada de las subespecies de S. salamandra en la Península Ibérica.
-S. s. almanzoris Müller y Hellmich, 1935: La caracterización morfológica y molecular de esta subespecie muestra la existencia de tres grupos de poblaciones en las Sierras de Gredos, Guadarrama y San Vicente (Figura 2) (Martínez-Solano et al., 2005). Se localiza generalmente por encima de los 1800 m, en prados subalpinos con lagunas de origen glaciar. Por su elevada actividad acuática (García-París, 1985) y pequeño tamaño, ha sido considerada un ecotipo de S. s. bejarae (Gasser, 1978), aunque pertenecen a linajes evolutivos independientes (Steinfartz et al., 2000; García-París et al., 2003; Martínez-Solano et al., 2005). Se caracteriza por presentar manchas pequeñas y amarillas, de contorno irregular y dispersas sobre el dorso, en el que predomina el color negro (Bosch y López-Bueis, 1994). Puede tener coloración rojiza en la cabeza y parótidas, aunque no es un carácter típico para la subespecie. Las poblaciones de S. s. almanzoris son ovovivíparas.
Figura 2. Salamandra de Peñalara(Madrid) (C). I. Martínez-Solano
-S. s. bejarae Wolterstorff, 1934: distribuida por todo el Sistema Central, Montes de Toledo, zonas montañosas de Cáceres y Badajoz, además de parte de Castilla y León. Se caracteriza por su forma rechoncha, morro puntiagudo, parótidas cortas y anchas, y dorso negro con un diseño de manchas amarillas e irregulares (Figura 3). En ocasiones presenta color rojizo en el cuerpo y/o parótidas. Tanto los análisis genéticos realizados hasta la fecha, como la propia distribución de esta especie (carente de un sentido biogeográfico) y su enorme semejanza con S. s. gallaica, apuntan a otro caso de polimorfismo dentro de S. salamandra y por lo tanto la validez de esta subespecie debería ser estudiada. Las poblaciones de S. s. bejarae son ovovivíparas.
Figura 3. Salamandra de Villablino (León) (C). G. Velo-Antón
-S. s. bernardezi Mertens y Müller, 1940: se distribuye por el norte de las provincias de A Coruña y Lugo, y en Asturias. Se asemeja morfológicamente a S. s. fastuosa, presentando un tamaño pequeño (hasta 180 mm; Velo-Antón et al., 2015b), diseño rayado, parótidas pequeñas y morro corto y ligeramente puntiagudo (Figuras 4 y 5), lo cual dificulta la distinción de ambas subespecies. Existen zonas de contacto con S. s. gallaica al norte de A Coruña y Lugo (Galán, 2007), y al este con S. s. fastuosa (García-París et al. 2003). Además, entra en contacto con S. s. bejarae a lo largo de la Cordillera Cantábrica. Raramente presenta manchas rojizas. En los valles de los ríos Tendi y Marea (Figura 6) (Piloña, Asturias) fue descrita la subespecie S. s. alfredschmidti (Köhler y Steinfartz, 2006) en base a la ausencia de líneas negras dorsolaterales propias de S. s. bernardezi y a las diferencias genéticas obtenidas en base a ADN mitocondrial (D-loop). Sin embargo, un estudio más reciente en que se amplió el área de estudio analizando el patrón de coloración de más de 1.100 individuos, e incluyendo análisis genéticos y de similitud de nicho ecológico, demostró que se trata de un ejemplo de polimorfismo dentro de S. s. bernardezi (Beukema et al., 2015). Por lo tanto, el patrón de coloración es también variable en esta subespecie y es común el color pardo o marrón oscuro en muchos individuos (Villanueva, 1993; Pasmans y Keller, 2000; Pasmans et al., 2004; Beukema, 2006). El viviparismo es el modo de reproducción común en esta subespecie, al menos en el centro y oeste de Asturias, aunque se observa también ovoviviparismo en las poblaciones gallegas y en el oriente asturiano. En las poblaciones urbanas de Oviedo se han observado una gran cantidad de partos, todos ellos vivíparos (Buckley et al., 2007; Álvarez, 2012; Velo-Antón et al., 2015b). Presenta un ciclo anual.
Figura 4. Salamandra de Arriondas (Asturias) (C). G. Velo-Antón
Figura 5. Salamandras de Navia (Asturias) (C). G. Velo-Antón
Figura 6. Salamandras del Valle de Marea (Asturias) (C). G. Velo-Antón
-S. s. crespoi Malkmus, 1983: La distribución de esta subespecie es muy reducida, localizándose entre los 200 y 600 m de altitud en la Sierra de Monchique, en el sudoeste del Alentejo y probablemente en el Algarve (Reis et al., 2011). Se caracteriza por su gran tamaño (hasta 250 mm) y por tener la cabeza pequeña y aplanada, el morro en punta, extremidades y dedos largos y diseño de manchas pequeñas en forma de punto o gancho en el dorso, pudiendo formar pequeñas líneas transversales. Puede presentar coloración rojiza, al igual que S. s. gallaica.
-S. s. fastuosa Schreiber, 1912: distribuida en los Pirineos centrales y occidentales, País Vasco y Cantabria, y estableciendo zonas de contacto al sur con S. s. bejarae (Veith et al., 1997), al oeste de Cantabria con S. s. bernardezi (García-París et al., 2003), y en el Pirineo central con S. s. terrestris. El dorso es rayado y el grosor de las líneas es bastante variable (Figuras 7 y 8), generalmente sin interrupciones hasta llegar a su distribución más oriental, Benasque y el Val d’Aran, donde sí se observan individuos con un rayado discontinuo (Montori y Herrero, 2004). No presenta manchas rojizas. Es una subespecie de tamaño pequeño, morro corto y redondeado, cola y dedos relativamente largos y parótidas pequeñas. Presenta las dos estrategias reproductoras, viviparismo y ovoviviparismo (Uotila et al., 2013) y un ciclo bianual en las poblaciones pirenaicas (Gasser y Joly, 1972).
Figura 7. Salamandra de Santander (C). G. Velo-Antón
Figura 8. Salamandras de San Sebastián (Guipúzcoa) (C). G. Velo-Antón
-S. s. gallaica Seoane, 1884: se distribuye por Galicia y Portugal, contactando con poblaciones de S. s. crespoi al sur de su rango de distribución (Malkmus, 1983), y con poblaciones de S. s. bernardezi al noreste (Bas, 1983; Galán, 2007) y S. s. bejarae al este. Los individuos de esta subespecie son por lo general de gran tamaño y hocico puntiagudo. El tamaño en las poblaciones gallegas oscila entre 120 y 200 mm (Galán y Fernández-Arias, 1993, Cordero et al., 2007; Velo-Antón et al., 2015b), pudiendo llegar hasta 250 mm (Ferrand de Almeida et al., 2001). Podría decirse que S. s. gallaica es la subespecie que muestra mayor variación en pigmentación y diseño (Figura 9). Aunque la combinación del negro y amarillo son los colores predominantes en este grupo, también es frecuente observar individuos melánicos, e individuos con un elevado porcentaje de color rojo y/o gris distribuido por todo el cuerpo (Guiberteau et al., 2012). Las poblaciones insulares de San Martiño y Ons tienen una marcada tendencia al melanismo, presentando menor proporción de amarillo, rojo y gris que las poblaciones costeras próximas a estas islas. En cuanto al patrón de manchas, también varían desde el típico patrón de manchas irregulares a diseños con líneas discontinuas. Las poblaciones de S. s. gallaica son ovovivíparas, exceptuando las poblaciones insulares de las Rías Baixas (Velo-Antón et al., 2007; 2012; 2015b) y la zona de contacto con S. s. bernardezi a lo largo del norte y el oriente gallego, donde son vivíparas (Bas, 1983; Galán, 2007). En los alrededores de Coimbra también se han detectado casos de viviparismo (Fachbach, 1976), aunque no se ha podido volver a demostrar.
Figura 9. Salamandras de los alrededores de Pontevedra (C). G. Velo-Antón
-S. s. longirostris Joger y Steinfartz, 1994: se localiza en la Sierra de Ronda, Sierra de Grazalema, al este del Guadalhorce y al oeste de Sierra Nevada. La longitud total alcanza los 230 mm y se caracteriza por tener un morro corto y muy puntiagudo (con forma de cuña desde una visión lateral), la cola corta y rechoncha, parótidas grandes de color amarillo, y un diseño dorsal con manchas cuadrangulares de color amarillo alineándose en dos bandas longitudinales (Figura 10). Presenta también manchas amarillas en la parte superior de los ojos y en la parte posterior de la boca. No presenta manchas de coloración rojiza. Este linaje subespecífico es el único que presenta una discontinuidad geográfica con el resto de distribución de la especie, con entidad morfológica propia y con un claro aislamiento genético a nivel mitocondrial (García-París et al., 1998, 2003), aunque todavía no se ha demostrado su aislamiento genético con marcadores nucleares. Sus poblaciones son ovovivíparas.
Figura 10. Salamandra de Cádiz (C). I. Martínez-Solano
-S. s. morenica Joger y Steinfartz, 1994: Se distribuye desde Sierra Morena a las Sierras de Segura, Cazorla y Alcaraz. La longitud total alcanza los 210 mm y se caracteriza por tener la cabeza redondeada, un morro puntiagudo y manchas moteadas en el dorso que en ocasiones forman líneas similares a S. s. crespoi. Las parótidas tienen coloración amarilla o roja (Figura 11).
Figura 11. Salamandra de la Sierra de Cazorla (Jaén) (C). J. Bosch
-S. s. terrestris Bonnaterre, 1789: en la península Ibérica esta subespecie se distribuye en el Pirineo oriental hasta el Val d’Aran, y se extiende por Cataluña a través de la Serralada Litoral y Prelitoral hasta la Serra del Montsant-Prades (Montori y Herrero, 2004; Donaire et al., 2014). Como ocurre con otras subespecies, muestra una elevada variedad de patrones de coloración dorsal, desde manchas amarillas formando dos líneas continuas, líneas formadas por manchas más irregulares y manchas situadas cerca de la línea vertebral (Rivera y Simón, 1999; Donaire et al., 2014). Fuera de la Península Ibérica se extiende por la mayor parte del territorio francés, centro y norte de Europa. Presenta una talla media, y morro algo redondeado. La estrategia reproductora es el ovoviviparismo y presenta ciclo anual.
Referencias Alcobendas, M., Dopazo, H., Alberch, P. (1994). Genetic structure and differentiation in Salamandra salamandra populations from the northern Iberian Peninsula. Mertensiella, 4: 7-23. Alcobendas, M., Dopazo, H., Alberch, P. (1996). Geographic variation in allozymes of populations of Salamandra salamandra (Amphibia: Urodela) exhibiting distinct reproductive modes. Journal of Evolutionary Biology, 9 (1): 83-102. Álvarez, D. (2012). Las salamandras de la ciudad de Oviedo: una vida entre el asfalto. Quercus, 321: 26-32. Balogová, M., Uhrin, M. (2015). Sex-biased dorsal spotted patterns in the fire salamander (Salamandra salamandra). Salamandra, 51 (1): 12-18. Barbadillo, L. J. (1987). La Guía de Incafo de los Anfibios y Reptiles de la Península Ibérica, Baleares y Canarias. Incafo, Madrid. Bas, S. (1983). Estudio de la situación microevolutiva y de la ecología de Salamandra salamandra (L.) en el Noroeste Ibérico. Tesis Doctoral. Universidad de Valencia. Bas, S., Gasser, F. (1994). Polytypism of Salamandra salamandra (L.) in North-western Iberia. Mertensiella, 4: 41-74. Benavides, J., Viedma, A., Clivilles, J., Ortiz, A., Gutiérrez, J. M. (2000). Albinismo en Alytes dickhilleni y Salamandra salamandra en la Sierra del Castril (Granada). Boletín de la Asociación Herpetológica Española, 11: 83. Beukema, W. (2006). Filling the gap in the distribution of Salamandra salamandra alfredschmidti Köhler & Steinfartz 2006, and remarks on the reproduction of the Rio Tendi vally salamanders in Asturias, Spain. Amphibia, 5 (2): 20-23. Beukema, W. (2011). Ontogenetic pattern change in amphibians: The case of Salamandra corsica. Acta Herpetologica, 6 (2): 169-174. Beukema, W., Nicieza. A. G., Lourenço, A., Velo-Antón, G. (2015). Colour polymorphism in Salamandra salamandra (Amphibia: Urodela), revealed by a lack of genetic and environmental differentiation between distinct phenotypes. Journal of Zoological Systematics and Evolutionary Research. En prensa. Boada, M., Aguirre, D., Maluquer-Margalef, J. (2012). Troballa d’un exemplar de Salamandra comuna Salamandra salamandra (Linnaeus, 1758) albina al Montnegre (Vallès Oriental; Catalunya). Butlletí de la Societat Catalana d'Herpetologia, 19: 180-183. Bosch, J., López-Bueis, I. (1994). Comparative study of the dorsal pattern in Salamandra salamandra bejarae (Wolterstorff, 1934) and S. s. almanzoris (Müller & Hellmich, 1935). Herpetological Journal, 4: 46-48. Buckley, D. (2003). La evolución del viviparismo en Salamandra salamandra (Linnaeus, 1758). Tesis Doctoral. Universidad Autónoma de Madrid. Buckley, D., Alcobendas, M., García-París, M., Wake, M. H. (2007). Heterochrony, cannibalism, and the evolution of viviparity in Salamandra salamandra. Evolution & Development, 9: 105-115. Campeny, R. (1991). Variaciones en las branquias de larvas de Salamandra salamandra en hábitats naturales hipóxicos. Revista Española de Herpetología, 6: 101-106. Cordero, A., Velo-Antón, G., Galán, P. (2007). Ecology of amphibians in small coastal Holocene islands: local adaptations and the effect of exotic tree plantations. Munibe, 25: 94–103. Díaz-Paniagua, C., López-Jurado, L. F. (1981). Notas sobre las larvas de urodelos de la mitad sur de la Península Ibérica: caracteres diferenciadores y desarrollo. Boletín de la Estación Central de Ecología, 10 (20): 27-34. Donaire, D., Rivera, X., Fernández, D. (2014). Colour pattern of Salamandra salamandra (Linnaeus, 1758) in Catalonia and Europe. Butlletí de la Societat Catalana d'Herpetologia, 21: 37-74. Dopazo, H. J., Boto, L. Alberch, P. (1998). Mitochondrial variability in viviparous and ovoviviparous populations of the urodele Salamandra salamandra. Journal of Evolutionary Biology, 11: 365-378. Eiselt, J. F. (1958). Der Feuersalamander, Salamandra salamandra, Beiträge zu einer taxonomischen Synthese. Abhandlungen und Berichte für Naturkunde und Vorgeschichte Museum Magdeburg, 10, 77-154. Escoriza, E., García-Cardenete, L. (2006). Polimelia en Alytes dickhilleni y Salamandra salamandra longirostris. Dos casos de ejemplares con seis extremidades. Boletín de la Asociación Herpetológica Española, 16 (1-2): 39-41. Fachbach, V. G. (1976). Biologie, Taxonomie und phylogenetische Beziehungen der verschiedenen Unterarten von Salamandra salamandra im Bereich der Iberischen Halbinsel. Journal of Zoological Systematics and Evolutionary Research, 14 (1): 59-78. Ferrand de Almeida, N., Ferreira, A., Teixeira, J. A., Catalão, A. I. (2001). Anfíbios e Répteis de Portugal. FAPAS, Porto. Frost, D. R. (2014). Amphibian Species of the World: an Online Reference. Version 6.0. http://research.amnh.org/herpetology/amphibia/index.html. American Museum of Natural History, New York, USA. Galán. P., Fernández Arias, G. (1993). Anfibios e Réptiles de Galicia. Edicións Xerais, Vigo. Galán. P. (2007). Viviparismo y distribución de Salamandra salamandra bernardezi en el norte de Galicia. Boletín de la Asociación Herpetológica Española, 18: 44-48. Galán, P., Velo-Antón, G., Cordero-Rivera, A. (2011). Puesta de huevos infecundos en Salamandra salamandra. Boletín de la Asociación Herpetológica Española, 22: 86-89. García-París, M. (1985). Los Anfibios de España. Ministerio de Agricultura, Pesca y Alimentación, Madrid. García-París, M., Alcobendas, M., Alberch, P. (1998). Influence of the Guadalquivir River Basin on the mitochondrial DNA evolution of Salamandra salamandra (Caudata: Salamandridae) from southern Spain. Copeia, 1998: 173-176. García-París, M., Alcobendas, M., Buckley, D., Wake, D. B. (2003). Dispersal of viviparity across contact zones in Iberian populations of fire salamanders (Salamandra) inferred from discordance of genetic and morphological traits. Evolution, 57: 129-143. Gasser, F. (1967). Differences au niveau des proteines sériques de type serumalbumine chez Salamandra salamandra terrestris et Salamandra salamandra fastuosa (Urodèle, Salamandridae). Comptes Rendues Hebdomadaires des Scéances de la Académie des Sciences série D, 265: 704. Gasser, F. (1978). Le polytypisme de l'espèce paléarctique Salamandra salamandra (L.) (Amphibien, Urodèle). I. Protéines sériques et groupes sériques. Archives de Zoologie Expérimentale et Générale, 119: 585-617. Gasser, F., Joly, J. (1972). Existence d’un cycle sexuel biennal chez la femelle de Salamandra salamandra fastuosa Schreiber (Urodèle, Salamandridae) a différentes altitudes dans les Pyrénées centrales: influence des facteurs génétiques et climatiques. Annales des Sciences Naturelles Zoologie et Biologie Animale, 14: 427-444. González de la Vega, J. P. (1988). Anfibios y Reptiles de la Provincia de Huelva. Ertisa, Huelva. Guiberteau, D. F., Vázquez Graña, R., López, J. E. (2012). Variabilidad de patrones y pigmentación en Salamandra salamandra gallaica. Butlletí de la Societat Catalana d'Herpetologia, 20: 115-120. Hillenius, D. (1968). Notes on Salamandra salamandra ssp. Bijdragen tot de Dierkunde, 38:31-38. Joger, U., Steinfartz, S. (1994). Zur subspezifischen gliederung der südiberischen feuersalamander (Salamandra salamandra-komplex). Abhandlungen und Berichte für Naturkunde, 17: 83-98. Köhler, G., Steinfartz, S. (2006). A new subspecies of the fire salamander, Salamandra salamandra (Linnaeus, 1758) from the Tendi valley, Asturias, Spain. Salamandra, 42: 13-20. Labus, N., Cvijanović, M., Vukov, T. (2013). Sexual size and shape dimorphism in Salamandra salamandra (Amphibia, Caudata, Salamandridae) from the central Balkans. Archives of Biological Sciences, 65 (3): 969-975. Lizana, M., Márquez, R., Martín-Sánchez, R., Ciudad, J., López, A., Orfao, A. (2000). Determination of cellular DNA content of Iberian salamanders by flow cytometry. Amphibia-Reptilia, 21 (4): 411-418. Llorente, G. A., Montori, A., Santos, X., Carretero, M.A. (1995). Atlas dels Amfibis i Rèptils de Catalunya i Andorra. El Brau, Barcelona. Malkmus, R. (1983). Beschreibung einer neuen Form des Feuersalamanders der Serra de Monchique (Portugal): Salamandra salamandra (gallaica) crespoi n. subsp. Faunistiche Abbandlungen, 10: 169-174. Mancino, G., Barsacchi, G., Nardi, I. (1969). The lampbrush chromosomes of Salamandra salamandra (L.) (Amphibia Urodela). Chromosoma, 26 (4): 365-387. Martínez, M. O., Fesser Justes, C. (2000). Los Anfibios del Alto Aragón. Instituto de Estudios Altoaragoneses, Huesca. Martínez-Solano, I., Alcobendas, M., Buckley, D., García-París, M. (2005). Molecular characterisation of the endangered Salamandra salamandra almanzoris (Caudata, Salamandridae). Annales Zoologici Fennici, 42: 57-68. Mertens, R., Müller, L. (1940). Liste der Amphibien und Reptilien Europas (2. Liste). — Abh. Senckenb. naturf. Ges., Frankfurt, 451: 1-56. Montori, A., Francesch, J. J. (1995). Morphometric analysis of Salamandra salamandra in the Iberian Peninsula: Preliminary data. Scientia Herpetologica, 1995: 37-40. Montori, A., Herrero, P. (2004). Caudata. En: García-París, M., Montori, A., Herrero, P. Amphibia, Lissamphibia. En: Ramos, M. A., et al. (Eds.). Fauna Ibérica, vol. 24. Museo Nacional de Ciencias Naturales, CSIC, Madrid. Müller, L., Hellmich, W. (1935). Mitteilungen über die Herpetofauna der Iberischen Halbinsel I. Über Salamandra salamandra almanzoris n. ssp. und Bufo bufo gredosicola n. ssp., zwei neue Amphibienrassen aus der Sierra de Gredos. Zoologischer Anzeiger, 112: 49-57. Palaus i Soler, X. (1999). Un cas de melanisme a Salamandra salamandra (L.). ). Butlletí de la Societat Catalana d'Herpetologia, 14: 95-96. Pasmans, F., Bogaerts, S., Keller, H. (2004). Note on the distribution of Salamandra salamandra cf. bernardezi in Asturias, northern Spain. Podarcis, 5: 58-60. Pasmans, F., Keller, H. (2000). Morphological variation in neighbouring populations of Salamandra salamandra bernardezi in northern Spain. Zeitschrift für Feldherpetologie, 7: 77-84. Ragghianti, M., Bucci-Innocenti, S., Mancino, G. (1982). C-banded karyotype and cytotaxonomy of Mertensiella caucasica (Waga, 1876)(Caudata: Salamandridae). Amphibia-Reptilia, 3 (4): 303-307. Reis, D. M., Cunha, R. L., Patrao, C., Rebelo, R., Castilho, R. (2011). Salamandra salamandra (Amphibia: Caudata: Salamandridae) in Portugal: not all black and yellow. Genetica, 139 (9): 1095-1105. Rivera, J., Simón, J. G. (1999). Diseños en la coloración dorsal en Salamandra salamandra (Linnaeus, 1758), de una población de Badalona (Catalunya). Butlletí de la Societat Catalana d'Herpetologia, 14: 70-77. Rivera, J., Simón, J. G., Vilagrosa, A., Fonoll, R. (1994). Datos sobre la presencia de salamandras (Salamandra salamandra L. 1758) de manchas rojo-anaranjadas en la Península Ibérica. Boletín de la Asociación Herpetológica Española, 5: 25-27. Rivera, X., Donaire-Barroso, D., Arribas, O. (2014). Hipótesis sobre el rigen y función del patrón de coloración y de la estrategias reproductivas en el género Salamandra Laurenti, 1768. Butlletí de la Societat Catalana d'Herpetologia, 21: 75-92. Salvador, A., García-París, M. (2001). Anfibios Españoles: Identificación, Historia Natural y Distribución. Esfagnos, Talavera de la Reina. Schreiber, E. (1912). Herpetologia Europaea. Eine systematische Bearbeitung der Amphibien und Reptilien welche bisher in Europa auggefunden sind. Verlag von Gustav Fischer, Jena. 960 pp. Seoane, V. L. (1884). Identidad de Lacerta schreiberi (Bedriaga) y Lacerta viridis, Var. Gadovii (Boulenger), e investigaciones herpetológicas de Galicia. Abad, La Coruña. 20pp. Steinfartz, S., Veith, M., Tautz, D. (2000). Mitochondrial sequence analysis of Salamandra taxa suggests old splits of major lineages and postglacial recolonizations of Central Europe from distinct source populations of Salamandra salamandra. Molecular Ecology, 9 (4): 397-410. Uotila, E., Díaz, A. C., Azkue, I. S., Rubio Pilarte, X. (2013). Variation in the reproductive strategies of "Salamandra salamandra" (Linnaeus, 1758) populations in the province of Gipuzkoa (Basque Country). Munibe Ciencias Naturales. Natur zientziak, 61: 91-101. Veith, M. (1997). Allozyme clines in Salamandra salamandra populations from the Cantabrian Mountains, Spain. Amphibia-Reptilia, 18: 298-303. Velo-Antón, G., Buckley, D., Daoudi, A. D., Rivera, A. C. (2007a). Bicephaly in Salamandra salamandra larvae. Herpetological Bulletin, 101: 31-32. Velo-Antón, G., Buckley, D., Martínez-Solano, I., Canestrelli, D., Lourenço, A., Martínez-Freiría, F. (2015a). Reconstructing the biogeographic history of geographically disjunct populations: the case of S. s. bernardezi and S. s. giglioli. 18th SEH (European Congress of Herpetology), 7-12 September 2015, Wrocław, Poland. Velo-Antón, G., García-París, M., Galán, P., Cordero Rivera, A. (2007b). The evolution of viviparity in Holocene islands: ecological adaptation versus phylogenetic descent along the transition from aquatic to terrestrial environments. Journal of Zoological Systematics and Evolutionary Research, 45: 345-352. Velo-Antón, G., Santos, X., Sanmartín-Villar, I., Cordero-Rivera, A., Buckley D. (2015b). Intraspecific variation in clutch size and maternal investment in pueriparous and larviparous Salamandra salamandra females. Evolutionary Ecology, 29: 185-204. Velo-Antón, G., Zamudio, K. R., Cordero-Rivera, A. (2012). Genetic drift and rapid evolution of viviparity in insular fire salamanders (Salamandra salamandra). Heredity, 108 (4): 410-418. Vences, M., Sanchez, E., Hauswaldt, J. S., Eikelmann, D., Rodriguez, A., Carranza, S., Donaire, D., Gehara, M., Helfer, V., Lotters, S., Werner, P., Schulz, S., Steinfartz, S. (2014). Nuclear and mitochondrial multilocus phylogeny and survey of alkaloid content in true salamanders of the genus Salamandra (Salamandridae). Molecular Phylogenetics and Evolution, 73: 208-216. Villanueva, A. (1993). Hallazgo de una nueva coloración de Salamandra salamandra bernardezi en Asturias. Boletín de la Asociación Herpetológica Española, 4: 14-15. Villanueva, A. (2007). Polimelia en un ejemplar de Salamandra salamandra en Asturias. Boletín de la Asociación Herpetológica Española, 18: 90-91. Wolterstorff, W. (1934). Über Salamandra maculosa molleri und eine neue Form, subsp. bejarae. Bl. Aquar.-Terrar.-Kunde, 45: 147-159.
Guillermo Velo-Antón1 y David Buckley2,3 2Departamento de Biodiversidad y Biología Evolutiva. Museo Nacional de Ciencias Naturales MNCN-CSIC. c/ José Gutiérrez Abascal 2, Madrid 28006, España 3Dirección actual: 1 Bailey Mews, Auckland Road. Cambridge CB5 8DR Fecha de publicación: 2-12-2015 Velo-Antón, G., Buckley, D. (2015). Salamandra común – Salamandra salamandra. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Martínez-Solano, I. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|
|
|
|
|
|