|
|
|
|
|
|
Para más información sobre cada apartado hacer click en:
|
Key words: European Common Frog, description, size, variation
Sinónimos y combinaciones Rana temporaria Linnaeus, 1758; Rana aquatica Linnaeus, 1758; Rana hyla Linnaeus, 1758; Rana muta Laurenti, 1768; Rana alpina Laurenti, 1768; Rana campanisona Laurenti, 1768; Rana rufa Lacépède, 1788; Rana atra Bonnaterre, 1789; Ranaria temporaria Rafinesque, 1814; Rana alpina Risso, 1826; Rana flaviventris Millet de la Turtaudière, 1828; Rana temporaria var. canigonensis Boubée, 1833; Rana glacialis Boubée, 1833; Rana scotica Bell, 1839; Rana platyrrhinus Steenstrup, 1846; Rana fusca Thomas, 1855; Rana temporaria var. typus Koch, 1872; Rana temporaria var. montanus Koch, 1872; Rana temporaria var. maximus Koch, 1872; Rana temporaria var. verrucosus Koch, 1872; Rana temporaria var. cinereus Koch, 1872; Rana temporaria var. gracilis Koch, 1872; Rana temporaria var. acutirostris Fatio, 1872; Rana temporaria var. obtusirostris Fatio, 1872; Rana temporaria var. platyrrhina Schreiber, 1875; Rana temporaria fusca Bedriaga, 1881; Rana fusca honnorati Herón-Royer, 1881; Rana muta var. obtusirostris Camerano, 1884; Rana muta var. acutirostris Camerano, 1884; Rana muta var. subconcolor Camerano, 1884; Rana muta var. flavo-maculata Camerano, 1884; Rana muta var. nigro-guttata Camerano, 1884; Rana muta var. atra Camerano, 1884; Rana fusca var. longipes Müller, 1885; Rana temporaria parvipalmata López-Seoane, 1885; Rana (Crotaphitis) fusca Schulze, 1891; Rana temporaria parvipalmata Boettger, 1892; Rana temporaria var. canigonica Belloc, 1893; Rana temporaria var. marmorata Werner, 1897; Rana temporaria var. nigromaculata Werner, 1897; Rana temporaria var. entzi Werner, 1897; Bufo campanisonus Boulenger, 1897; Hyla hyla Prazák, 1898; Rana esculenta var. alpina Prazák, 1898; Rana muta platyrrhina var. concolor Prazák, 1898; Rana muta platyrrhina var. sordida Prazák, 1898; Rana muta platyrrhina var. albostriata Prazák, 1898; Rana muta platyrrhina var. paradoxa Prazák, 1898; Rana muta platyrrhina var. quaturolineata Prazák, 1898; Rana muta platyrrhina var. alpina Prazák, 1898; Rana méhelyi Bolkay, 1912; Rana temporaria subsp. mehelyi Ehik, 1921; Rana muta muta Mertens, 1925; Alytes campanisonus campanisonus Lindholm, 1929; Alytes grandis Brunner, 1957; Rana temporaria honnorati Mertens and Wermuth, 1960; Rana temporaria canigonensis Dubois, 1983; Rana (Rana) temporaria Dubois, 1987; Rana honnorati Mensi, Lattes, Macario, Salvidio, Giacoma, and Balletto, 1992; Rana aragonensis Palanca-Soler, Vieites, and Martínez-Suárez, 1995; Rana temporaria aragonensis Veith, Kosuch, and Vences, 2003; Rana temporaria canigonensis Veith, Kosuch, and Vences, 2003; Rana temporaria parvipalmata Veith, Kosuch, and Vences, 2003; Rana temporaria honnorati Veith, Kosuch, and Vences, 2003; Rana (Laurasiarana) temporaria Hillis and Wilcox, 2005 (Frost, 2008; García-París, 2000).
Descripción del adulto Rana grande y de aspecto robusto. Presenta dos manchas características a ambos lados de la cabeza que son más oscuras que el resto del cuerpo y que adquieren la forma de un antifaz. Estas manchas se extienden desde las narinas hasta las axilas, atravesando el ojo y el tímpano. El hocico es redondeado, los ojos tienen la pupila horizontal y el iris tiene reflejos dorados con algunas manchas parduzcas. El tímpano es grande, muy marcado y con un tamaño 2/3 menor que el ojo. En las patas anteriores, el tercer dedo de la mano es siempre mayor que el resto. Las patas posteriores son más cortas y rechonchas que en otras ranas pardas. Cuando se estira a lo largo del cuerpo, la articulación tibio-tarsal llega al ojo, pero no alcanza el extremo del hocico, lo que sirve como rasgo diagnóstico para diferenciarla de otras especies, como la Rana patilarga. Las membranas interdigitales no son muy grandes y normalmente solo cubren entre 1/2 y 2/3 de la longitud de los dedos. Los dedos tienen nódulos desarrollados, en las patas anteriores presentan tres tubérculos palmares y las patas traseras presentan tubérculos subarticulares en todos los dedos y un tubérculo metatarsal grande en la parte interna que mide entre 1/3 y 1/2 del primer dedo. La piel es lisa aunque las hembras la tienen más áspera, pudiendo presentar gránulos en su superficie. Tiene los pliegues dorsolaterales casi paralelos y próximos entre sí. La coloración en esta especie es muy variable, pudiendo encontrarse ejemplares completamente negros, grises, naranjas o incluso rojos. Asimismo presentan un patrón de manchas muy variado, desde un diseño completamente liso a ejemplares jaspeados y manchados. Muchos ejemplares presentan una mancha en forma de V invertida en la espalda. La coloración del vientre es más clara que el dorso, con matices blancos o amarillentos, pudiendo aparecer manchas de extensión variable (García-París, 1985, 2000; Salvador y García-París, 2001; Gollmann et al., 20141). La coloración dorsal negra aumenta con la edad en poblaciones de montaña de Pirineos (Riobo et al., 1999). Se ha encontrado en el Parque Natural das Fragas do Eume un ejemplar sin extremidad anterior izquierda (Galán, 2011).
Dimorfismo sexual Las hembras alcanzan un mayor tamaño que los machos y su aspecto es más redondeado, sobre todo antes de realizar la puesta. Los machos tienen sacos vocales internos y sus miembros anteriores son más robustos y musculados que los de las hembras. Durante la época de celo, los machos presentan unas callosidades negruzcas en los pulgares que ayudan al macho a sujetarse a la hembra durante el amplexus. La piel de la hembra es más áspera que la de los machos.
Tamaño La Rana bermeja es la mayor de las ranas pardas ibéricas, llegando algunas hembras grandes a alcanzar los 90 mm de longitud total (García-París, 1985) superando esta longitud en algunas poblaciones leonesas (Diego-Rasilla y Ortiz-Santaliestra, 2009). En los muestreos realizados por nuestro grupo de trabajo en la cordillera cantábrica, el ejemplar de mayor tamaño que encontramos fue una hembra de 105 mm de longitud en unas charcas temporales cerca del Cuetu Arbás (Cangas del Narcea) en 1999. Salvador y García-París (2001) hablan de tamaños máximos para esta especie de 111 mm para los machos y 107 mm para las hembras. Tal como se comenta en el apartado de Variación geográfica, existen importantes diferencias de tamaño a lo largo del gradiente altitudinal, siendo considerablemente más grandes los ejemplares de alta montaña que los que habitan en zonas bajas.
Descripción de los huevos y de la larva Las puestas tienen forma globosa y en ocasiones pueden alcanzar más de 20 cm de diámetro. Aunque nada más poner los huevos la puesta se hunde, al poco tiempo la gelatina que rodea a los huevos empieza a hidratarse y la puesta acaba subiendo a la superficie. En ocasiones la hembra no pone todos los huevos de una vez y se pueden observar varias pequeñas masas de huevos procedentes de una misma hembra. Los huevos son de color negro y miden entre 1,5 y 3 mm. El número de huevos que pone cada hembra depende de su tamaño, pero lo normal es que una puesta se componga de entre 1.000 y 2.000 huevos, aunque en algunos casos se han confirmado puestas de hasta 4000 huevos. Las larvas miden entre 6 y 8 mm al nacer. Al liberarse de la gelatina del huevo son de un color gris oscuro o negro que se va aclarando a medida que crecen, adquiriendo una coloración parda y ligeramente jaspeada con matices dorados y cobrizos. La cola es larga y acabada en punta y tiene una longitud de 3 a 4 veces mayor que su altura. La apertura anal se sitúa en el lado derecho y el espiráculo en el lado izquierdo. La parte ventral es oscura por lo que el paquete intestinal no se transparenta a través de ella (Galán, 1982; Salvador y García-París, 2001).
Características citogenéticas y bioquímicas Los estudios de electroforesis de proteínas realizados en varias poblaciones ibéricas indican que la especie presenta una elevada variabilidad interpoblacional, con distancias genéticas relativamente altas, sobre todo entre R. temporaria parvipalmata y R. t. temporaria (Arano et al., 1993). La Rana bermeja posee 26 pares de cromosomas, de características similares a los de R. dalmatina y R. iberica, con las regiones NOR situadas en el brazo largo del par 10, aunque el patrón de bandas C es diferente al de las dos especies anteriores. Asimismo se han encontrado diferencias en este patrón entre individuos ibéricos y los individuos alemanes (Herrero et al., 1990).
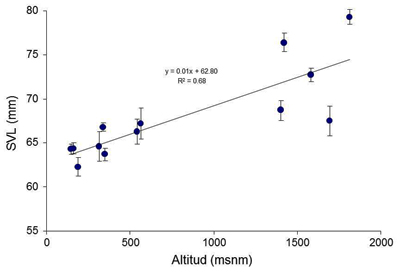
Variación geográfica En esta especie hay una gran variabilidad, tanto inter como intrapoblacional. Taxonómicamente en la Península ibérica se encuentran dos subespecies, R. t. parvipalmata Seoane, 1885, que se distribuye por toda la cornisa cantábrica, desde Galicia hasta Cantabria y parte del País Vasco, incluyendo el norte de León y el norte de Zamora, y R. t. temporaria Linnaeus, 1758, que se distribuye desde la parte oriental del País Vasco, hasta el Pirineo. Las dos subespecies son muy similares en su aspecto externo, si bien R. t. parvipalmata es de menor tamaño que la subespecie nominal (Veith et al., 2002, 2003). En el noroeste ibérico los machos miden de media 48,4 mm y las hembras 52,2 mm. También en esta subespecie es menor la extensión de la palmeadura interdigital (Galán, P. (1989a). El canto de R. t. parvipalmata se compone de menos impulsos que el de R. t. temporaria (Vences, 1992). Existen diferencias de tamaño a lo largo del rango altitudinal, siendo mayores a medida que se asciende en altitud. En Asturias a lo largo de los últimos años hemos realizado muestreos exhaustivos a lo largo de todo el gradiente altitudinal y observamos que a medida que ascendía el tamaño de los machos solía ser mayor y además mostraban acúmulos de grasa más desarrollados, sobre todo en aquellas poblaciones que pasaban un periodo de hibernación bajo la nieve (Figura 1) (David Álvarez y Alfredo G. Nicieza, datos no publicados).
Figura 1. Longitud media (± se) de los machos de 11 poblaciones asturianas de Rana temporaria y relación con la altitud de sus poblaciones.
Hay dos grupos de poblaciones que se correponden con los Pirineos por el este y Galicia y Asturias por el oeste; las poblaciones intermedias muestran distintos grados de introgresión en los genomas mitocondrial y nuclear (Veith et al., 2012).
Aunque las poblaciones de alta montaña en Pirineos son morfológicamente distintas de las poblaciones de baja altitud, no forman grupos genéticamente diferenciados (Veith et al., 2012). Las poblaciones aisladas del sur de Galicia, incluidas las de Manzaneda-Queixa y las de Trevinca-Segundeira, presentan escasa divergencia mitocondrial respecto de las poblaciones del norte, lo que sugiere que la colonización de las zonas meridionales es relativamente reciente y que ha mantenido el flujo de genes durante periodos más fríos (Galán et al., 2010). Los haplotipos del este y del oeste coexisten en España, Suiza y Francia, sugiriendo que ha habido expansión del clado oriental a través de un corredor mediterráneo (Teacher et al., 2009).
Referencias Arano, B., Esteban, M., Herrero, P. (1993). Evolutionary divergence of the Iberian brown frogs. Annales des Sciences Naturelles Zoologie 14: 49-57. Diego-Rasilla, F. J., Ortiz-Santaliestra, M.E. (2009). Los Anfibios. Serie Naturaleza en Castilla y León. Asociación Herpetológica Española. Servicio de Publicaciones de la Caja de Burgos. 237 pp. Frost, D.R. (2008). Amphibian Species of the World: an Online Reference. Version 5.2 (15 July, 2008). American Museum of Natural History, New York, USA. Galán, P. (1982). Biología de la reproducción de Rana iberica Boulenger, 1879 en zonas simpátridas con Rana temporaria Linneo, 1758. Doñana Acta Vertebrata 9: 85-98. Galán, P. (1989a). Diferenciación morfológica y selección de hábitats en las ranas pardas del noroeste ibérico, Rana iberica Boulenger, 1879 y Rana temporaria parvipalmata Seoane, 1885. Treballs de la Societat Catalana de Ictiología i Herpetología, 2: 193-209. Galán, P. (2011). Anfibios con malformaciones en el Parque Natural das Fragas do Eume (A Coruña, Galicia). Boletín de la Asociació Herpetológica Española, 22: 65-67. Galán, P., Ludewig, A.K., Kmiec, J., Hauswaldt, S., Cabana, M., Ferreiro, R., Vences, M. (2010). Low mitochondrial divergence of rediscovered southern relict populations of Rana temporaria parvipalmata in Spain. Amphibia-Reptilia, 31 (1): 144-148. García-París, M. (1985). Los anfibios de España. Publicaciones de Extensión Agraria. Madrid. García-París, M. (2000). Anura. Pp. 275-480. En: García-París, M., Montori, A., Herrero, P. (Eds.). Amphibia. Lissamphibia. En: Ramos, M. A. et al. (Eds.). Fauna Ibérica. Vol. 24. Museo Nacional de Ciencias Naturales, CSIC, Madrid. 639 pp. Gollmann, B., Borkin, L., Grossenbacher, K., Weddeling, K. (2014). Rana temporaria Linnaeus 1758- Grassfrosch. Pp. 304-437. En: Grossenbacher, K. (Ed.). Handbuch der Reptilien und Amphibien Europas. Band 5/III A. Froschlurche (Anura) III A (Ranidae I). Aula Verlag, Wiebelsheim. Herrero, P., Arano, B., Esteban, M. (1990). Karyotypic characterization of brown frogs from the Iberian Peninsula (Ranidae: Rana). Pp. 138-140. En: Cytogenetics of Amphibia and Reptilia. Olmo (Ed.). Birkhausen Verlag. Basel. Riobo, A., Rey, J., Puente, M., Miramontes, C., Vences, M. (1999). Ontogenetic increase of black dorsal pattern in Rana temporaria. British Herpetological Society Bulletin, 70: 1-6. Salvador, A., García-París, M. (2001). Anfibios españoles. Identificación, historia natural y distribución. Canseco Editores. Talavera de la Reina. Teacher, A.G.F., Garner, T.W.J., Nichols, R.A. (2009). European phylogeography of the common frog (Rana temporaria): routes of postglacial colonization into the British Isles, and evidence for an Irish glacial refugium. Heredity, 102 (5): 490-496. Vences, M. (1992). Zur Biologie der nordwestspanischen Braunfrosche Rana iberica Boulenger, 1879 und Rana temporaria parvipalmata Seoane, 1885. Salamandra, 28 (1): 61-71. Veith, M., Vences, M., Rodríguez Vieites, D., Nieto Román, S., Palanca-Soler, A. (2002). Genetic differentiation and population structure within the Spanish Rana temporaria complex (Amphibia: Ranidae). Folia Zool., 51: 307-318. Veith, M., Kosuch, J., Vences, M. (2003). Climatic oscillations triggered post-Messinian speciation of Western Palearctic brown frogs (Amphibia, Anura, Ranidae). Mol. Phyl. Evol.,26: 310-327.
David Álvarez Fecha de publicación: 8-05-2013 Otras contribuciones: 1. Alfredo Salvador. 27-10-2014 Álvarez, D. (2014). Rana bermeja – Rana temporaria. En: Enciclopedia Virtual de los Vertebrados Españoles. Salvador, A., Martínez-Solano, I. (Eds.). Museo Nacional de Ciencias Naturales, Madrid. http://www.vertebradosibericos.org/
|
|
|
|
|
|
|